|
Siberian Forest Insects: Ready for Exports
Yuri N. Baranchikov - Institute of Forestry, Siberian Branch, Russian Academy of Science, Krasnoyarsk 660036, Russia.
From: Exotic Pests of Eastern Forests, Conference Proceedings - April 8-10, 1997, Nashville, TN, Edited by: Kerry O. Britton, USDA Forest Service & TN Exotic Pest Plant Council
Introduction
Existing publications on Palaearctic insect invaders to North America
forests are devoted exclusively to Europe-U.S. comparisons (Niemela, Mattson,
1996). This is understandable from both geographical and historical points
of view. But as the history of mankind continues, new economical relations
are established which, unfortunately, make new possibilities for the introductions
of pests. The recent discovery in New York of a cerambycid beetle Anoplophora
glabripennis native to Asia (USDA Forest Service, 1996) and the well
known "Asian Gypsy Moth Case" (Wallner, 1996) are the reminders
that the forests of Northern and Northeastern Asia are an important source
of exotic pests. The largest part of this region belongs to the Russian
Federation.
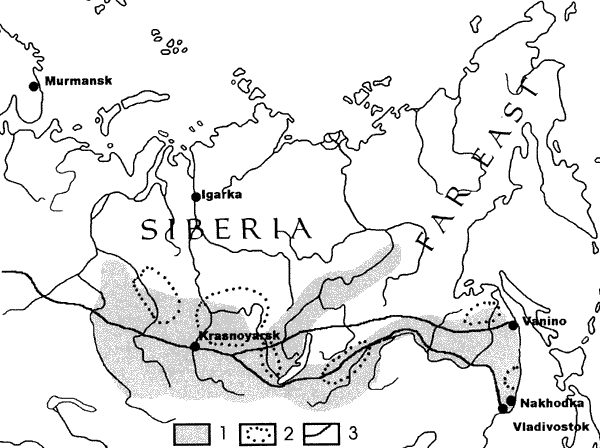
The forested territory of Asian Russia could be devided
in two parts (Fig. 1): Siberia (from Urals to the Khabarovsk Kray) and the
Far East (with administrative units of Khabarovsk and Prymorsky Kray, Amurskaya,
Magadanskaya, Kamchatskaya and Sakhalinskaya Oblast'). The forest land of
this region spans a wide range of latitudes, elevations, precipitation,
and soils. More than 60 percent of forests in Asian Russia grow on the permafrost
(Pozdniakov, 1986).
Comparison of the forest composition of Siberia and Northern North America
shows that they are similar in type, but differ in species composition.
In Siberia 81% of the forested territory is covered with conifers. Nearly
all of the conifer forest is composed of six species: Larix |
 Figure 1. Forest insect injury zones and
areas of logging and transportation in
Siberia and the Russian Far East. 1 – area
of most severe outbreaks; 2 – primary areas
of logging; 3 – Trans-Siberian railroad. |
sibrica,
Larix dahurica, Pinus silvestris, P. sibirica, Abies
sibirica and Picea obovata. Larch forests dominate both in area
(62%) and in growing stock (52%) (Falaleev, 1985).
Even a brief comparison shows the similarity of Asian Russian forests
to that of Northwestern North America. In the Western U.S., 82% of forests
are conifers (Powell et al., 1993), but the list of the woody species is
slightly longer than that of Siberia and the Russian Far East. Though Siberia
and Northern U.S. and Canada are dissimilar at the species level of woody
plants, these regions are much more similar on the genus level. There are
at least 16 genera of trees found in both areas (e.g. Abies, Alnus, Betula,
Colylus, Crataegus, Juniperus, Larix, Picea, Pinus, Populus, Rhamnus, Salix,
Sorbus, Tilia, Ulmus, and Viburnum). Taxonomic diversity of these genera
in Siberia is comparable with those in the different regions of the Northern
U.S. and Canada (Table 1).
Table 1. Number of tree species per woody plant genus in Siberia
and Northern United States and Canada (Data from Koropachinskiy, 1983; Elias,
1980; introduced species not included).
| Plant Genera |
Siberia |
Alaska |
Western Canada |
Northwestern U.S. |
Eastern Canada |
Northeastern U.S. |
Northern North America |
| Abies |
1 |
– |
3 |
5 |
1 |
1
| 8 |
| Alnus |
5 |
2 |
4 |
4 |
1 |
3
| 7 |
| Betula |
8 |
1 |
2 |
2 |
5 |
6
| 7 |
| Cornus |
2 |
1 |
2 |
3 |
2 |
4
| 6 |
| Craetagus |
4 |
– |
2 |
2 |
6 |
11
| 12 |
| Juniperus |
5 |
1 |
2 |
3 |
1 |
1
| 4 |
| Larix |
3 |
1 |
2 |
3 |
1 |
1
| 3 |
| Picea |
2 |
3 |
4 |
3 |
3 |
3
| 7 |
| Pinus |
3 |
– |
6 |
9 |
2 |
8
| 17 |
| Populus |
6 |
3 |
3 |
4 |
4 |
5
| 8 |
| Prunus (Padus) |
1 |
– |
3 |
3 |
4 |
8
| 10 |
| Rhamnus |
4 |
– |
1 |
2 |
– |
2
| 3 |
| Salix |
11 |
8 |
14 |
13 |
10 |
12
| 25 |
| Sambucus |
1 |
– |
1 |
1 |
2 |
2
| 4 |
| Tilia |
1 |
– |
– |
– |
1 |
2
| 2 |
| Ulmus |
4 |
– |
– |
– |
3 |
5
| 5 |
| Euonymus |
2 |
– |
– |
1 |
– |
1
| 2 |
The taxonomic diversity of forest insects in Asian Russia
is comparable with those of boreal zones of Northern America. For example,
218 and 212 species of bark beetles (Scolytidae) were reported from Asian
Russia and Canada respectively (Danks, 1979; Yanovskiy, 1996).
Only 90 species of insects are of real economical importance in the forests
of Siberia and the Far East (Baranchikov, Montgomery, 1996). The main folivores
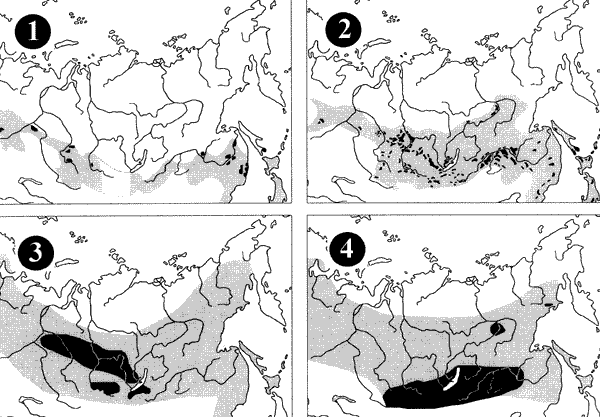
are: Dendrolimus superans, Lymantria dispar and Zeiraphera
grizeana on Larix; Lymantria monacha (Fig. 2) and Bupalis
pinearius on Pinus; D.superans on Abies and Choristoneura
murinana on Picea. The major wood borers are: Ips cembrae
and Xylotrechus altaicus on Larch; Ips sexdentatus and Tomicus
piniperda on Pinus; Monochamus urussovi on Abies;
and Ips typographus on Picea. The following three species
are the most widespread and destructive:
Fir sawyer beetle (Monochamus urussovi Fisch.)
is a transpalearctic species occuring in coniferous forests from Finland
to the Pacific Ocean (Fig. 2). The insect infests nearly all species of
Pinaceae but the firs (Abies) are most heavily damaged. The beetle
vectors the phytopathogenic fungus Ceratocystis sp. During their
feeding on the crown, the adult beetles remove strips of bark and infect
branches with the fungus spores. The developing fungus kills tiny branches
on the periphery of the crown, weakens the tree and reduces resin flow.
This makes oviposition and larvae development of the beetle more successful.
To our knowledge fungus is pathogenic only for fir species. In Siberia,
M. urussovi is frequently found on birches (Betula), but causes
little damage to it. |
 Figure 2. Distribution (grey) and areas of economic importance
(black) of some forest
insect pests of Northern Asia: 1 – Lymantria
monacha; 2 – Dendrolimus superans; 3 – Monochamus
urussovi; 4 – Ips cembrae. |
The life cycle of M. urussovi typically lasts for 2 years. The
beetles fly from late May or early June through September. A female lays
eggs under the bark, one at a time; eggs hatch in 16 to 30 days. From the
second instar and up to pupation the larvae gnaw tunnels in the wood. Winter
is usually spent in the larval stage. Before pupation, larvae form pupal
chambers in the wood, separated from the surface by a thin layer of bark
and wood where they pupate. The pupal phase lasts from 4 to 5 weeks; adults
emerge by gnawing a round hole 6 to 12 mm in diameter through the bark.
M. urussovi is one of the most destructive pests of firs in Northern
Asia. The pest increases its number in Abies forests damaged by defoliating
insects, fires and windfalls. Having infested the damaged parts of the forest,
the beetle population becomes dense enough to attack, weaken and kill healthy
stands. By attacking healthy fir stands dense beetle populations can maintain
outbreak levels indefinitely, causing the death of fir forests over enormous
areas. In the late 1950's in the Tomsk Oblast' (Western Siberia), the pest
destroyed 2 million m3 of fir wood stock. This caused the collapse of forest
enterprises in that region for years and previously planned construction
of a railway was delayed significantly slowing the industrial development
of the whle area. In 1971-1976, an outbreak of M. urussovi destroyed
300,000 hectares of fir forest in Krasnoyarsk Kray in the Central Siberia
(Isaev et a., 1988).
Larch bark beetle (Ips cembrae Heer) is known
in Russian literature as Ips subelongatus Motschulsky. It is a transpalearctic
species occurring from Europe to Japan (Fig. 2). In Siberia it attacks all
species of larch, spruce (Picea) and pine, but larch is the principal
host on which outbreaks occur.
The pest has 2 generations per year in the southern regions of Siberia
where the yearly sum of temperatures above 10 C% is more than 1500 and the
frost-free period lasts more than 2.5 months. The adult beetles hibernate,
mostly in the litter, then emerge from late May to early June , attack trees
and lay eggs. Larval and pupal stages occur from June to early July. First
generation beetles emerge and attack trees in July. The larval and pupal
stages occur from late July to mid-August. New adults start emerging in
mid-August. They feed on the same tree if the previous density was not too
high, or migrate to neighboring trees. In Europe, I. cembrae adults have
been recorded feeding in tree crowns, much like the bark beetles of the
genus Tomicus. Such crown feeding has not been recorded in Siberia.
In Northern Asia, I. cembrae is particularly destructive in the
larch forests of the south taiga forest- steppe complex. The distinguishing
features of these stands are enhanced aridity and high temperatures in summer,
the conditions under which I. cembrae can produce two generations. When
the forest is damaged by factors that kill more than 20% of the trees, the
bark beetle can become epidemic and outbreaks occur. During the outbreak
it can infest resistant larch forests adjacent to the damaged ones, thus
making the losses much greater.
Siberian moth (Dendrolimus superans sibiricus
Tschtvrk.) is widely distributed in Urals, Siberia, and the Far East (Fig.
2). Outbreaks occur in Abies sibirica, Pinus sibirica, Picea
spp. and Larix spp. forests, although larvae feed on most conifers
in the family Pinaceae.
The length of the life cycle varies from two to four calendar years depending
on population density. The larvae of the males have 5 to 9 instars, those
of the females 6 to 10; typically males have 5 and females 6. The larvae
are up to 110 mm long. Moths fly from the end of June to the beginning of
August and lay eggs on needles or branches. Commonly two winters are spent
in the larval stage; second to third instars and fifth to sixth instars
overwinter coiled up, under the forest litter. Pupation occurs from mid-June
to late July in cocoons in tree crowns. During outbreaks, a large portion
of excessively dense populations has a life cycle of two calendar years
and the rest have a three year cycle. As a result, the adults of two generations
emerge simultaneously and the population increases sharply. At the depression
phase, some portion of the population have a four calendar year life cycle,
where three winters are spent as larvae.
D. superans is the major defoliator of coniferous forests in Asian
Russia. In the fir-dominated forests of Central Siberia there were 10 outbreaks
since 1873, the last 5 were carefuly documented. They occured in 1935-1947,
1950-1959, 1962-1969, 1978-1985 and 1989-1997 defoliating 0.7, 2.6, 0.9,
0.1 and 1.1 million ha respectively. These forests all died, either directly
from the defoliation or from the increasing attacks of the fir sawyer beetle
or fire. In the South Siberia, D. superans outbreaks take place in
larch forests. Outbreaks on larch are not as destructive as those on firs
because larch is very tolerant to defoliation.
The ways of possible introductions of Siberian forest
insects are through the sea ports of Asian Russia. The overall dry cargo
shipments through all Russian ports in the year to July 1996 were 34 million
tonnes of which 24 million were exports. The largest Russian port is Novorossyisk
on the Black sea, handling 16% of all Russian cargo. The northern port of
Murmansk represents 8% and combined Far Eastern ports (Nakhodka, Vladivostok,
Vostochny and Vanino) represent the largest concentration at 31% of the
total Russian capacity. Large amount of cargo shipments, a deficit of manpower,
and the complex economic situation in Russia make the task of the Russian
Far East Quarantine Service extremely difficult (Gordon, 1996). International
cooperation should be enhanced to prevent transmision of exotic organisms.
The mutual Russian-U.S. project on monitoring the population level of lymantriid
moths in the Far Eastern port areas was started in 1993 (U.S. Department
of Agriculture, 1996). It was agreed that insect outbreaks in the nearby
forests should trigger mitigation measures.
Trains transport 97 % of all cargo that enters and leaves the port area.
The main transport artery of Asian Russia is the Trans-Siberian railroad
(Fig. 1). For much of its length, the railroad goes through the areas known
as "the zone of forest insect injury" - the area of the most severe
outbreaks and forest damage (Epova and Pleshanov, 1996). Open railroad cars
with wood and containers are exposed to many kinds of natural infestations
during the 2-6 weeks that it takes them to go through Siberia. Currently,
lymantriid pest populations are monitored at the Far Eastern ports area,
but this activity is not enough. On the vast area from Urals to Pacific
Ocean, flying gypsy moth females can freely put eggs on the containers at
the railway stations which are brightly illuminated at night.To more efficiently
prevent the occurrence of pest insects on cargo we need the entomological
information from all of the Siberian zone of potential infestation, as well
as port areas.
Mutual efforts between Russia and U.S.A were set on
technology and information transfer regarding pest risk assessment and control
of potential pests. Besides the lymantriid survey project, mentioned above,
efforts were made to access existing scientific information on species of
risk and on the methods of their control (U.S.Department of Agriculture,
1991; Wallner et al., 1995; Baranchikov et al., 1996; Baranchikov and Montgomery,
in prep.).
Literature Cited
Baranchikov, Y.N., Montgomery, M.E. (Eds.) (In prep.). Major insect forest
pests of Northern Asia. (In preparation).
Baranchikov, Y.N., Nikitenko G.N., Montgomery, M.E. 1996. The Russian
and Ukrainian literature on the gypsy moth, p. 48. In: Proceedings, U.S.Department
of Agriculture Interagency gypsy moth research forum 1995. U.S. Forest Service
Gen.Tech. Report NE-213, Radnor, PA: U.S. Forest Service.
Danks, H.V. (Ed.). 1979. Canada and its insect fauna. Memoirs of the
Entomological Society of Canada, 108, 573 p.
Elias, T.S. 1980. Trees of North America. The complete field guide and
natural history. New York: Van Nostrand Reinhold Co. 948 p.
Epova, V.I.; Pleshanov, A.S. 1996. Zony vredonosnosti nasekomykh-fitofagov
asiatskoi chasti Rossii [Phytophagous insects injury zones in the Asiatic
Russia]. Novosibirsk: Nauka Publ. 46 p. (In Russian).
Falaleev, E.N. 1985. Lesa Sibiri [Siberian forests]. Krasnoyarsk: KGU
Publ. 134 p.(In Russian).
Gordon, D. 1996. Enforcement in the "Wild East": problems implementing
APHIS' new regulations, pp. 78-80. In: Importing wood products: pest risk
to domestic industries. Portland, OR: Oregon State University.
Isaev, A.S.; Rozkov, A.S.; Kiselev, V.V. 1988. Chorny pihtovy usach'
Monochamus urussovi Fisch. [Fir sawyer beetle Monochamus urussovi Fisch.].
Novosibirsk: Nauka Publ. 270 p. (In Russian).
Koropachinskiy, I.Y. 1983. Drevenye rastenia Sibiri [Woody plants of
Siberia]. Novosibirsk: Nauka Publ. 383 p. (In Russian).
Niemela, P.; Mattson, W. 1996. Invasion of North American forests by
European phytophagous insects. BioScience. Vol. 46, 1: 741-753.
Powell, D.S.; Faulkner, J.L.; Darr, D.R.; Zhiliang Zhu; MacCleery, D.W.
1993. Forest resources of the United States, 1992. Gen. Tech. Rep. RM-234.
U.S.Department of Agriculture, Forest Service. 25 p.
Pozdniakov, L.K. 1986. Merzlotnoe lesovedenie [Forestry on the permafrost].
Novosibirsk: Nauka Publ. 191 p. (In Russian).
USDA Forest Service. 1996. Asian cerambycid beetle. A new introduction.
Durham, NH: USDA Forest Service, Forest Health Protection. 1 p.
U.S. Department of Agriculture. 1991. Pest risk assessment of the importation
of larch from Siberia and the Soviet Far East. Miscellaneous Publications
No. 1495. Washington, DC: U.S. Forest Service.
U.S. Department of Agriculture. 1996. Russian Far East Lymantriid monitoring
project. Project summary 1995. FHP Report 96-03. Ogden, Utah: U.S.Forest
Service.
Wallner, W.E. 1996. Invasion of the tree snatchers. American Nurseryman,
March:28-30.
Wallner, W.E., Humble, L.M., Levin, R.E., Baranchikov, Y.N., Carde, R.T.
1995. Response of adult Lymantriid moths to illumination devices in the
Russian Far East. Journal of Economic Entomology 88: 337-342.
Yanovskiy, V.M., 1996. Annotirovannyi spisok koroedov (Coleoptera, Scolytidae)
Severnoi Azii [Annotated list of Scolytidae of the Northern Asia]. Krasnoyarsk:
Institute of Forest Russian Academy of Sciences. 52 p. (In Russian).
|