|
|
In: Van Driesche, R., et al., 2002, Biological Control of Invasive Plants in the Eastern United States, USDA Forest Service Publication FHTET-2002-04, 413 p.
noxious weed laws of Florida (FDEP, 2000), Louisiana (LDWF, 2000), Texas (TPWD, 2000), California (CDFA, 2000a), South Carolina (SCDNR, 2000), North Carolina (NCAWCA, 2000), Oregon (OSDA, 2000), Washington (WSDA, 2000), and Arizona (ERDC 2001b). In addition, the states of Alabama, Georgia, Maryland, Mississippi, Tennessee, and Virginia, have programs for the control of this invasive plant (Eubanks, 1987; Earhart, 1988; Zattau, 1988; Bates, 1989; Henderson, 1995; Center et al., 1997).
resulting in $2,650,000 of expenses due to repairs, dredging, and fish loss. In addition, during this repair period, there was an estimated $2,000,000 loss in power generation for the plant (letter from Charleston District Engineer to Commander, South Atlantic Division, dated March 8, 1993).
Boat marinas have been reported closed for extended periods on the Potomac River, Virginia; Lake Okeechobee, Florida; Santee Cooper Reservoirs, South Carolina; and Clear Lake, California. Propeller driven boats are hampered by thick mats of hydrilla that form at the water’s surface, requiring frequent cleaning to progress short distances. The fragmented plant material removed from the propellers can easily colonize new areas. In the late 1980s, hydrilla populations at Lake Guntersville, Alabama increased rapidly. Henderson (1995) examined the economic impact of aquatic plant control programs on recreational use of this lake between 1990 and 1994. He found that the greatest economic value for recreation ($122 million annually) occurred when vegetation levels were 20% of the total lake area, and that revenue declined as hydrilla acreage increased. Although California does not consider hydrilla established, the state has, for decades, aggressively pursued an eradication program that seeks to rapidly eliminate new infestations as they are discovered. California officials have stated that if infestations are not contained and treated promptly, hydrilla will spread throughout the state and cost millions of dollars annually to manage (CDFA, 2000b). Ecological damage. Native plants act as the primary producers in most ecosystems (Drake et al., 1989; Pimm, 1991). In the United States, hydrilla frequently forms large monocultures that displace native vegetation (Haller, 1978), reducing biodiversity and altering native ecosystems. These alterations also affect the primary and secondary consumers in affected communities (Westman, 1990; Frankel et al., 1995; Schmitz and Simberloff, 1997). Massive amounts of hydrilla can alter dissolved oxygen, pH, and other water chemistry parameters (Smart and Barko, 1988). The portion of the water column occupied by aquatic plants also influences the presence and size distribution of fish (Killgore et al., 1993; Harrel et al., 2001). In dense hydrilla mats, feeding by certain predatory fish is hampered, and small insectivores predominate, reducing community diversity. (Dibble et al.,1996). Extent of losses. Hydrilla is a major aquatic weed problem throughout the southeastern United States (Center et al., 1997). It was introduced to North America in 1951 or 1952 by an aquarium plant dealer who discarded six bundles of hydrilla into a canal near his business in Tampa, Florida (Schmitz et al., 1991). Since then, it has spread explosively because it can reproduce from very small fragments (Langeland and Sutton, 1980). Apparently, recreational boaters and fishermen quickly spread hydrilla to new locations when fragments of hydrilla are transported on boats, motors, and trailers. Once an aquatic site is infested, eradication of hydrilla is very difficult. It produces specialized asexual, reproductive ‘buds’ on stems (referred to as turions) and on the underground stolons (tubers). These tubers and turions assist hydrilla in reinfesting a site after a drought, or after application of herbicides. Langeland (1990) reported that the annual control cost to manage 7,600 ha of hydrilla in Florida exceeds $5 million. The U.S. Army Corps of Engineers spends more than one million dollars per year to suppress hydrilla populations in the Jacksonville District and more than $400,000 annually to treat infestations of this plant at Lake Seminole, a 30,000-acre lake located on the borders of Florida, Alabama, and Georgia. Since 1989, millions of dollars have been spent to introduce the triploid grass carp into the Santee Cooper Reservoirs (70,000 ha) for the management of more than 17,000 ha of hydrilla (Morrow et al., 1997; Kirk et al., 1996, Kirk et al., 2000). Grass carp populations have reduced the infestation levels of hydrilla; however, additional stocking may be needed to maintain the current level of control (Kirk et al., 2000), which will also add to the management costs of this program. Hydrilla was first reported in California in 1976, and at that time the state established an eradication management plan. This program has eradicated hydrilla from various sites in ten counties. At some sites, treatment of hydrilla continued for six to eight years before eradication was achieved. Funding for this program has gradually increased over time, and during the last three years, California has spent more than $5.39 million (nearly $1.8 million annually) to eradicate hydrilla infestations in that state (CDFA, 2000a). Geographical Distribution Hydrilla is now almost cosmopolitan in its distribution. Antarctica and South America are the only continents from which it has not been recorded. It is very common on the Indian subcontinent, many of the Middle East countries, Southeast Asia, and northern and eastern Australia. Based on C. D. K. Cook’s (pers. comm.) list of herbarium specimens, hydrilla is found in the Southern Hemisphere as far south as the North Island of New Zealand (approximately 40° S). In the Northern Hemisphere hydrilla is found as far north as Ireland, England, Poland, Lithuania and Siberia. The Lithuanian sites, at about 55° N latitude, are the furthest from the equator that hydrilla is known to occur. Since virtually the entire continental United States, except Alaska, lies below a latitude of 48°, hydrilla is climactically suited for growth in any of the contiguous states as well as Hawaii. Even Alaska cannot be considered entirely safe from invasion by hydrilla since places such as Juneau are at approximately the same latitude as the hydrilla infestations in Lithuania and Siberia (Balciunas and Chen, 1993). The female form of dioecious hydrilla arrived in Florida in the early 1950s (Schmitz et al., 1991) and quickly spread throughout the southeastern United States. Although the monecious biotype of hydrilla was not detected in the United States until the late 1970s (Haller, 1982; Steward et al., 1984), it too is now spreading rapidly, especially into northern states. Monecious hydrilla has now been detected as far north as the Columbia River in Washington state in the western United States, and in Pennsylvania and Connecticut in the eastern United States (Madeira et al., 2000). An excellent color map showing the current U.S. distribution of both biotypes of hydrilla can be found in Madeira et al. (2000).
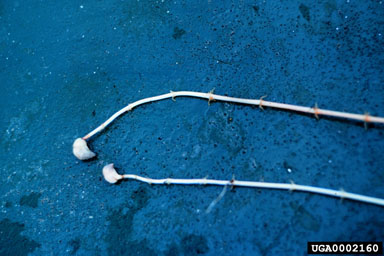
plants remain viable and are a common mode for infestation of new areas. Below the hydrosoil, the stems are horizontal, creeping, and stoloniferous. Leaves are verticillate, and along most of the stem, usually number three to five per node. Apical portions of the stem usually have the nodes tightly clustered, with each verticil bearing up to eight leaves. The leaves are usually strongly serrated with the teeth visible to the naked eye, and each leaf terminates in a small spine. The midvein is sometimes reddish in color, and is usually armed with an irregular row of spines. The squamulae intravaginales (nodal scales) are small (ca. 0.5 mm long), paired structures at the base of the leaves and are lanceolate, hyaline, and densely fringed with orange-brown, finger-like structures called fimbrae. Flowers are imperfect (unisexual), solitary, and enclosed in spathes. The female flower is white, translucent, with three broadly ovate petals, about 1.2 to 3.0 mm long; the three petals alternate with the sepals that are much narrower and slightly shorter; the three stigmas are minute; the ovary is at the base of a long (1.5 to 10+ cm) hypanthium. The male flower is solitary in leaf axils. Mature flowers abscise and rise to the surface. Sepals and petals are similar in size and shape to those of female flowers. Each of three stamens bears a four-celled anther that produces copious, minute, spherical pollen. Hydrilla plants occur as two biotypes. They can be either dioecious, with flowers of only one sex being produced on a particular plant, or monecious, with flowers of both sexes on the same plant. Fruits are cylindrical, about 5 to 10 mm long, usually with long, spine-like processes. Seeds are smooth, brown, usually five or less, 2 to 3 mm long and borne in a single linear sequence. Two types of hibernacula are produced – a brown, bulb like type is produced at the ends of the stolons (Fig. 3), while a green, conical form is found in axils of branches. In the United States, the first type is usually called tubers and the latter turions.
Biology Although the female biotype of hydrilla quickly became widespread throughout the southeastern United States, it was not until 1976 that a male flower was observed in the United States (Vandiver et al., 1982). The female flowers can only be pollinated in the air. The female flower reaches the water surface by elongation of the hypanthium (flower “stalk”). The petals and sepals of the female flower form an inverted bell with an air bubble when growing to the surface, and if after reaching the surface the flower becomes submerged, the petal and sepals revert to this position, and enclose an air bubble thus preventing wetting of the stigmas and ensuring air pollination. The male flower lacks a hypanthium, and reaches the surface by detaching from the plant and floating up as a ripe, air-filled bud. The perianth segments recurve towards the water surface and eventually the anthers dehisce, explosively scattering pollen in a radius of about 10 cm around the flower. Where male hydrilla flowers are present, the water surface frequently becomes visibly greenish-white due to the floating pollen grains and discarded male flowers. Hydrilla is usually a gregarious plant that frequently forms dense, intertwined mats at the water’s surface. Approximately 20% of the plant’s biomass is concentrated in the upper 10 cm of such a mat (Haller and Sutton, 1975). The plants grow and spread quickly. Small fragments of the plant, containing but a single node, can quickly develop adventitious roots and eventually produce an entire plant. Hydrilla has very wide ecological amplitude, growing in a variety of aquatic habitats. It is usually found in shallow waters, 0.5 m or greater in depth. In very clear waters it can grow at depths exceeding 10 m. It tolerates moderate salinity – up to 33 percent of seawater (Mahler, 1979). While hydrilla flourishes best in calcareous ponds and streams, water quality rarely seems to be limiting, since it is found in both acidic and alkaline waters. It also grows well in both oligotrophic and eutrophic waters, and even tolerates high levels of raw sewage (Cook and Lüönd, 1982). Sediments with high organic content provide the best growth, although hydrilla also is found growing in sandy and rocky substrates. While hydrilla does not grow well in deeply shaded areas, it is adapted to grow under very low light conditions (Bowes et al., 1977), and this may account for its rapid growth and quick dominance over native vegetation. Analysis of Related Native Plants in the Eastern United States While hydrilla can assume widely different forms when growing in different environments, all are now considered to be a single species of Hydrilla verticillata (Cook and Lüönd, 1982). There are no other species in the genus Hydrilla, which is placed in the frog’s bit family, Hydrocharitacae. There are eight other genera from this family in the eastern United States, two of which (Halophila and Thalassia) are native “marine grasses” that grow in shallow coastal waters (Godfrey and Wooten, 1979). The other native Hydrocharitacae, all of which grow in shallow freshwaters, include Blyxa aubertii Rich., Elodea (two species, Elodea canadensis Michaux and Elodea nutallii [Planch.] St. John), Limnobium spongia (Bosc.) Steud., and Vallisneria americana Michx. (Godfrey and Wooten, 1979). There also are three additional introduced Hydrocharitaceae in the United States: Egeria densa Planch. Hydrocharis morus-ranae L., and Ottelia alismoides (L.) Pers. The two native Elodea species, and the introduced Egeria densa, are difficult to distinguish readily from hydrilla. Hydrilla, however, is unique in having nodal scales (squamulae intravaginales) and specialized, asexual reproductive organs – tubers and turions. History of Biological Control Efforts in the Eastern United States Area of Origin of Weed The area of origin of Hydrilla verticillata is not clear, but appears to be a broad region encompassing a large part of the Eastern Hemisphere and adjacent areas. Cook and Lüönd (1982), along with many other botanists, indicate that “its centre of origin lies in the warmer regions of Asia.” However, hydrilla has been in central Africa for a long time – it was collected by Speke during his 1860 to 1863 expedition to find the sources of the Nile (Speke, 1864) – and some botanists believe that it originated there (Tarver, 1978). Mahler (1979) is even more precise, stating “...with a center of distribution or origin in southeastern Uganda and northwestern Tanzania.” Hydrilla is also considered by some to be native to Australia (Sainty and Jacobs, 1981). The first records from Australia are from the early nineteenth century, soon after the arrival of European settlers. A recent DNA analysis of hydrilla collections from around the world (Madeira et al., 1997) supports the hypothesis of multiple introductions into the United States. The authors found that dioecious samples from the southern United States are more closely aligned with those from the Indian subcontinent, while the monoecious samples most closely resembled those from South Korea. Domestic Surveys and Natural Enemies Found Prior to initiating a biological control project, it is recommended that the target weed be surveyed to determine what natural enemies are already associated with it in the invaded area. Native insects or pathogens might be suppressing a target weed at some sites, or non-native natural enemies may have been introduced accidentally. The Army Corps of Engineers Waterways Experiment Station funded thorough faunistic surveys of U.S. hydrilla populations by University of Florida entomologist, Joe Balciunas. Between 1978 and 1980, he made 289 collections of hydrilla at 75 sites, 58 of which were in Florida (Balciunas and Minno, 1984). More than 17,000 insect specimens, comprising nearly 200 species, were collected and identified (Balciunas and Minno, 1984), but of these only 15 were feeding on hydrilla (Balciunas and Minno, 1985). Among the most damaging of the insects found in Florida was the introduced Asian moth Parapoynx diminutalis Snellen. This moth was first detected in south Florida (Delfosse et al., 1976), but dispersed rapidly to additional areas, at some of which it caused heavy damage to hydrilla (Balciunas and Habeck, 1981). Other researchers (Cuda et al., 1999, 2002; Epler et al., 2000) have commented on the feasibility of using the midge Cricotopus lebetis Sublette (Diptera: Chironomidae) as a biological control agent for hydrilla. The feasibility of using native pathogens to control hydrilla also has been investigated. In the fall of 1987 and 1988, surveys were conducted in 15 lakes and 3 rivers in southeastern United States for pathogens of hydrilla (Joye and Cofrancesco, 1991). Nearly 200 fungal and 27 bacterial isolates were collected from hydrilla foliage. An endemic fungal pathogen originally identified as Macrophomina phaseolina (Tassi) Goid. and later determined to be Mycoleptodiscus terrestris (Gerd.) Ostazeski was collected from hydrilla growing in Lake Houston, Texas in 1987 (Joye, 1990; Shearer, 1996). Field and laboratory studies have shown that the fungus can significantly reduce hydrilla biomass after inoculation compared with untreated plants (Joye, 1990; Shearer, 1996). Disease symptoms appear in 5 to 7 days after inoculation as interveinal chlorosis followed by a complete loss of color. Within 10 to 14 days, plants treated with M. terrestris begin to disintegrate (Joye, 1990; Shearer, 1996). Transmission electron microscopy studies have shown that the fungus attaches to lower epidermal cells of hydrilla leaves within eight hours postinoculation and penetration through the cell wall is completed within 40 hours (Joye and Paul, 1992). The fungus then completely colonizes the host, resulting in collapse of the entire plant. While not currently available as a product, M. terrestris is undergoing evaluation for its potential as a bioherbicide for hydrilla management. As an initial step in the process, the U.S. Army Engineer Research and Development Center Environmental Laboratory (ERDC), Vicksburg, Mississippi and the USDA, ARS National Center for Agricultural Utilization Research in Peoria, Illinois are studying fermentation methods that will yield high concentrations of effective propagules at a low cost. SePro Inc. (Carmel, Indiana) also is involved as a cooperator in the project. The goal is to produce a bioherbicide that can be competitive with chemical herbicides. Overseas Areas Surveyed and Natural Enemies Found Determining the native range of a weed is extremely important in biological control programs since the center of origin is usually considered to be the best area to begin searches for natural enemies. In its native range, the weed should have a greater array of natural enemies that coevolved with it. Since evidence to pinpoint hydrilla’s evolutionary origin was lacking, searches have been made in several regions, including Africa, Asia, and Australia. Opportunistic surveys began in India in 1968, and since that time surveys have been conducted in at least 15 additional countries. A time-line and list of overseas research to develop biocontrol agents for hydrilla is presented in Table 1. Only the major overseas projects will be discussed here, as it is beyond the scope of this chapter to completely review the results of all the surveys noted in Table 1. For a more complete review of the history of foreign exploration for hydrilla agents, readers should consult Balciunas (1985), Buckingham (1994), and Balciunas et al., (1996a). Table 1. Chronology of foreign searches for insect enemies of hydrilla (Hydrilla verticillata)
Many of the overseas surveys consisted of either brief trips to one or more countries, or efforts in which hydrilla was added as a target to a larger, ongoing project in a specific region. While these opportunistic surveys frequently noted potential agents, as of 2000, none of these had been approved or released in the United States. The most productive overseas studies have been intensive, multi-year projects concentrating on hydrilla natural enemies in a particular region. The first of these was the USDA-sponsored project in Pakistan from 1971 to 1976, conducted by scientists from CIBC (Commonwealth Institute of Biological Control). Ten insects were studied (Baloch and Sana-Ullah, 1974), but only three were recommended for importation into the United States (Baloch et al., 1980). Unfortunately, these recommendations were not acted upon, possibly because there was no USDA scientist or facility available at that time to work on hydrilla insects.
Table 2. Candidate biological control agents evaluated for use against hydrilla.
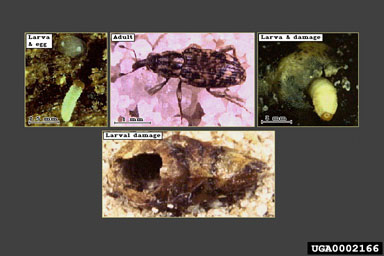
Between 1985 and 1992, Balciunas and his Australian staff made more than 100 non-quantitative collections and 588 quantitative collections of hydrilla at 70 sites in Australia (Balciunas et al., 1996a). In order to ascertain the field host range of the potential agents, he and his team also made 1,007 quantitative collections of 47 other aquatic plant species from 27 families (Balciunas et al., 1996a). Balciunas and his team evaluated six insects for their potential as biological control agents for hydrilla. Four of these were exported to the Florida quarantine for further evaluation (Table 2), and two were eventually released. In 1989, Balciunas joined USDA, ARS, and for three years headed a project, based at the Sino-American Biological Control Laboratory (SABCL), to find new agents for both hydrilla and Eurasian milfoil, Myriophyllum spicatum L., in temperate parts of China. Since then, the USDA, ARS Invasive Plant Laboratory in Ft. Lauderdale, Florida has led the searches in China for hydrilla natural enemies, and has expanded the surveys to Thailand and Vietnam (Table 1). Staff of this laboratory have been assisted in these surveys not only by SABCL scientists, but by other scientists from the United States and the Australian Biological Control Laboratory (ABCL). The most promising insects identified during the past decade are listed in Table 2. Overseas pathogens for controlling hydrilla also have been investigated, but far less extensively than the insects. During a three-month period in 1971 and 1972, surveys were conducted in India for pathogens of hydrilla (Charudattan, 1973). Of 40 fungi and 15 bacteria isolated and screened for pathogenicity, only two species, a Pythium sp. and a Sclerotium sp., were found to be damaging. Charudattan et al. (1980) reported that a pathogen, Fusarium roseum (Link ex Fr.) var. culmorum Snyd. and Hans. found on diseased Stratiotes aloides L. in The Netherlands, was efficacious on hydrilla. Staff of the Sino American Biological Control Laboratory also conducted surveys in the People’s Republic of China in 1994 and 1995 for pathogens of hydrilla. All isolates were subsequently deposited at the USDA, ARS quarantine facility located at Fort Detrick, Frederick, Maryland. Following identification of the isolates, they were subjected to pathogenicity screening on the host. Six isolates (an unidentified Moniliaceous hyphomycete, an unidentified Coelomycete, Phoma sp., Colletotrichum gloeosporioides [Penz.] Penz. and Sacc. in Penz., and M. terrestris were found to induce disease symptoms on hydrilla. Additional pathogenicity testing on rooted plants has yet to be completed. If potential biological control candidates are found among the isolates they will have to undergo intense host specificity testing because some have been reported on other hosts (Farr et al., 1989). Host Range Tests and Results The host range tests on the more than two dozen non-U.S. species of insects or pathogens that have been considered as potential biological control agents for hydrilla have been recorded in more than a hundred (mostly unpublished) reports. In Table 2, we summarize the primary test results for these potential agents. Only a few agents were tested extensively overseas, and their host range tests subsequently published in refereed journals, e.g., Balciunas and Center (1981), Balciunas and Burrows (1996), and Balciunas et al. (1996b). Nearly 20 hydrilla insect species were shipped to the quarantine facility in Gainesville, Florida for evaluation (Table 2). The testing there was conducted by Gary Buckingham, USDA, ARS, and University of Florida cooperators. Heightened concern for safety has increased the number of plant species tested, and the hydrilla agents eventually approved for release were tested on more than 60 species of plants in 30 families (Buckingham, 1994). Although a few species were conclusively ruled out as having too broad a host range, testing of many remains incomplete. Eventually, however, sufficient laboratory and field data was gathered to gain approval for release of two weevils and two leaf-mining flies. Although none of these four insects were strictly monophagous, hydrilla was greatly preferred, and the risk to the few other alternate hosts was considered very minimal. Releases Made Many of the natural enemies identified during overseas surveys still have not been fully evaluated to judge their safety as potential biological control agents for hydrilla. Only four hydrilla insects have been released in the United States: The tuber attacking weevil Bagous affinis Hustache (Coleoptera: Curculionidae) and the leaf mining fly Hydrellia pakistanae Deonier (Diptera: Ephydridae) were both released in 1987; another leaf-mining fly H. balciunasi Bock (Diptera: Ephydridae) was released in 1989; and the stem-mining weevil B. hydrillae O’Brien (Coleoptera: Curculionidae) was released in 1991 (Buckingham, 1994). The leaf-mining flies have been the most extensively released species. Hydrellia pakistanae has been released at more than 50 sites in Alabama, California, Florida, Georgia, Louisiana, and Texas (Center et al., 1997). About 1.2 million individuals were obtained, mainly from greenhouse colonies maintained at the U.S. Army Engineer Research and Development Center in Vicksburg, Mississippi and various USDA, ARS facilities, along with an additional two million insects from a Tennessee Valley Authority pond-based rearing facility (Grodowitz and Snoddy, 1995). These releases ended in 1995. Recently (September 2000), releases resumed using Hydrellia-containing hydrilla obtained from ponds at the Lewisville Aquatic Ecosystem Research Facility, Lewisville, Texas with more than 300,000 immatures being released in Lake Raven in Huntsville State Park, Texas. Although considerably less effort went into the release of H. balciunasi, still close to one million individuals were released at 11 sites in Florida and Texas only (Grodowitz et al., 1997). Bagous affinis was extremely difficult to maintain under mass-rearing conditions. This was due primarily to the high demand of tubers for larval feeding. However, over 10,000 individuals were released in three states (i.e., California, Florida, and Texas) at more than 10 locations (Godfrey et al., 1994; Grodowitz et al., 1995). A larger effort went into the release of the stem-feeding weevil, B. hydrillae. For example, close to 300,000 individuals have been released in four states (Florida, Texas, Georgia, and California) at more than 15 locations (Grodowitz et al., 1995). No overseas pathogens have yet been approved for release to control hydrilla. Biology and Ecology of Key Natural Enemies Hydrellia pakistanae - “Asian Hydrilla Leaf Mining Fly” and Hydrellia balciunasi - “Australian Hydrilla Leaf Mining Fly” (Diptera: Ephydridae)
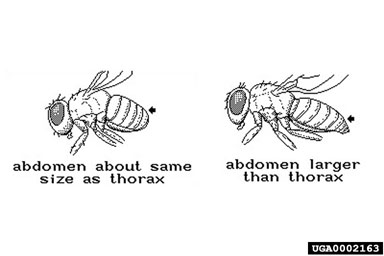
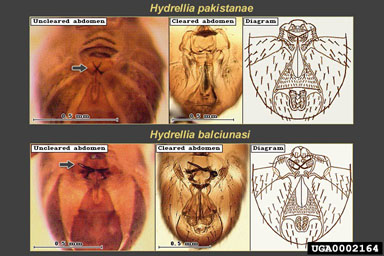
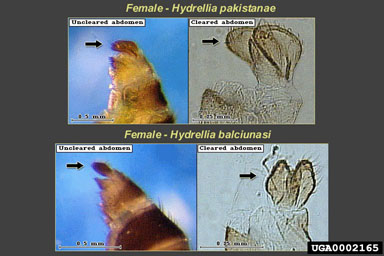
Adult H. pakistanae and H. balciunasi, the two introduced Hydrellia spp. can be difficult to identify because of their small size, lack of obvious distinguishing characters, and similarity to other native species of Hydrellia (including H. bilobifera Cresson and H. discursa Deonier). Examinations of reproductive organs are frequently required for positive identification. Adult male H. pakistanae can be distinguished from other commonly collected native Hydrellia spp. and H. balciunasi by several characters, including the length of the thorax in comparison to the abdomen length, the presence of
development time is from 20 to 35 days. The overwintering stage is unknown but larvae have been found on hydrilla throughout the entire winter. The total number of generations per growing season appears to be highly variable and related to geographic area but may be as high as seven.
From a distance, a hydrilla mat containing large numbers of Hydrellia spp. appears brown, and upon close examination, one can observe clusters of leaves along the stem where feeding has occurred. Damage to hydrilla is probably due to a reduction in total photosynthetic area caused by the leaf damage (Doyle et al., 2002), which reduces growth and vigor and leads to a decrease in the competitiveness of the affected plants. In addition, some evidence suggests that feeding may reduce the buoyancy of the plant and allow the stem to become more brittle in areas of heavy feeding, leading to stem fragmentation (Grodowitz et al., 1999). Limited field observations suggest that Hydrellia feeding may predispose the plant to infection by fungi and other pathogens. Bagous affinis - “hydrilla tuber weevil” (Coleoptera: Curculionidae)
There are three larval instars and they are non-descript and typically creamy-white. The larvae can be found on or within the hydrilla tubers, where they burrow and feed. The larvae pupate within the tubers but also can pupate in nearby moist wood. The duration of the larval stage is anywhere from 14 to 17 days. The pupal stage lasts four to six days.
While the adults feed on the tubers, their damage is minimal compared to the destructiveness of the larvae. The larvae can attack and destroy tubers deep within the sediment. High weevil populations have been reported from hydrilla-infested ponds in the insect’s native range. Bagous hydrillae - “hydrilla stem weevil” (Coleoptera: Curculionidae)
hydrilla, apparently preferring the stem tissue at the internodes. Eggs are laid within stem tissues usually at the leaf nodes. Eggs hatch in three to four days and larvae feed throughout internal stem tissues. Larval feeding subsequently fragments the stem, which floats to the shoreline where the third instars exit and subsequently pupate within soil or drying hydrilla. Pupation must take place under relatively dry conditions. The pupal period lasts from three to four days depending on the ambient temperature.
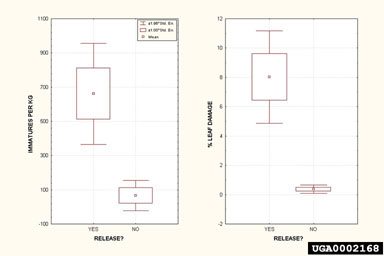
Since no permanent populations of B. hydrillae exist in the United States, large-scale damage has not been observed; however, researchers in Australia have indicated that larval feeding by B. hydrillae causes the plants to have a mowed appearance due to the removal of the hydrilla from the surface to a depth of 100 cm (Balciunas and Purcell, 1991). Evaluation of Project Outcomes Establishment and Spread of Agents Although four insects have been released, neither of the weevils appears to have established, and H. balciunasi has only been recovered from a few sites in east Texas (Bennett and Buckingham, 1999; Grodowitz et al., 2000a). However, H. pakistanae established and dispersed readily and is now found throughout Florida; north to Muscle Shoals, Alabama; west to Austin, Texas; and south to the lower Rio Grande Valley (Center et al., 1997; Grodowitz et al., 1997; Grodowitz et al., 2000a). Populations of both species, but especially H. pakistanae, have expanded in distribution considerably since they were first released. For example, H. pakistanae was released in the early 1990s at only one location, Lake Boeuf in extreme southern Louisiana, but surveys conducted in 2000 revealed its presence at several locations up to 300 km west and north of the original introductions (Freedman and Grodowitz, unpub.). In Florida, H. pakistanae is found associated with a majority of sites containing hydrilla infestations, indicating considerable range expansion (Center, 1992; Center, pers. comm.). In Texas, populations of H. pakistanae and H. balciunasi also have increased considerably from the four original release sites. One of the most interesting findings has been the discovery of H. pakistanae in the extreme south central portion of Texas on the Rio Grande, more than 250 km from the nearest release site (Grodowitz et al., 1999). Such range extensions are surprising since the introduced Hydrellia spp. are relatively weak fliers with short adult life spans. In addition, the non-contiguous lake systems in both Texas and Louisiana should have hampered range extension for these species. At many sites throughout the country, especially non-release sites, Hydrellia spp. population levels appear minimal with less than 200 immature insects/kg wet weight of hydrilla and leaf damage not exceeding 2%. Bagous affinis was originally described from India and Pakistan and was first released in the United States in Florida in 1987 (Bennett and Buckingham, 1991). As of the spring of 2001, no permanent populations were known to exist in the United States. Because of its strict environmental requirement for distinct wet/dry periods to allow access to buried tubers, this species has not been released at many sites. Releases in California, at locations where water levels can be controlled, have indicated that this weevil can successfully establish and, with appropriate water level management, overwinter (Godfrey et al., 1996). Unfortunately, because of the hydrilla eradication program in California, the hydrilla at the California site was destroyed soon after verifying overwintering. The use of biological control in conjunction with an aggressive eradication program is counterproductive. Bagous hydrillae was first released in the United States in Florida in 1991 (Grodowitz et al., 1995), but no established populations have been confirmed. Extensive surveys were initiated, however, no weevils have been recovered at actual release sites even after extended periods. Bagous hydrillae adults have only been collected after suspension of releases at one site, Choke Canyon Reservoir, Texas during 1993 and 1994 (Grodowitz et al., 1995). However, soon after the termination of releases B. hydrillae adults were no longer observed at Choke Canyon Reservoir. Suppression of Target Weed and Recovery of Native Plant Communities Impact of the introduced Hydrellia spp. has apparently been observed at several release sites in Georgia, Florida, and Texas. For example, significant changes have been observed in the hydrilla status at Lake Seminole, Georgia, over the last few years, following the release of more than 1.5 million H. pakistanae in 1992 (Grodowitz et al., 1995; Grodowitz, Cofrancesco, Stewart, and Madsen, unpub.). For the first several years following this large release, numbers of H. pakistanae in Lake Seminole remained at low but detectable levels based on the presence of immatures on randomly selected stem pieces and Berlese funnel extraction of plant material. Beginning in 1997, hydrilla populations began to decline in various areas of the lake and increases in plant diversity were observed that appeared related to increasing H. pakistanae populations. In 1999, large numbers of H. pakistanae adults were observed throughout large areas of the lake and these correlated with significant decreases in hydrilla populations and increases in other native plants, including several species of Potamogeton and Najas. Quantitative sampling of Hydrellia immatures based on stem counts and quantification of number of leaves damaged in September 1999 revealed the presence of more than 2,000 immatures per kg wet weight of hydrilla and close to 20% of the total number of leaves damaged. Quantitative plant sampling conducted during November showed significant reductions (ca. four-fold) in tuber numbers and three-fold increases in species richness in areas significantly affected by H. pakistanae feeding as observed in September. While reductions in tuber numbers were surprising, such reductions have been substantiated during large-scale, long-term replicated tank studies conducted recently (Grodowitz et al., 2000b; Doyle, Grodowitz, Smart, Owens, unpub.) and in short-term small container studies (Doyle et al., 2002). In these studies, lower number of tubers and biomass occurred in biological control treatments where damage exceeded 40% of the leaves only for short durations. Similar reductions in hydrilla were observed at Coleto Creek Reservoir, Texas in 1999 and 2000. Reductions in hydrilla were first observed in the two original release sites in 1998 continuing through 1999. Currently, only small quantities of hydrilla persist at the original release sites and reductions in hydrilla have been observed in a nearby cove where fly densities and hydrilla status were quantified for many years to be used as a control. In 1999, higher fly levels were observed in the control cove followed by substantial hydrilla declines in 2000. Observations of the lake in 2000 have shown increasing fly numbers and associated damage throughout the entire reservoir. Sampling of stems during November 2000 demonstrated leaf damages in the 12 to 15 % range for hydrilla located in the extreme northern portion of the lake (Grodowitz et al., 1999; Grodowitz, unpub.). Similar effects also have been observed in Sheldon Reservoir near Houston, Texas (Grodowitz et al., 1999). In these situations, as the hydrilla declined, it was apparently replaced with a mixture of submersed plants, including Eurasian watermilfoil (Myriophyllum spicatum L.), star grass (Heteranthera dubia [Jacquin] MacM.), coontail (Ceratophyllum demersum L.), and various species of Potamogeton and Najas, as well as an emergent species, American lotus (Nelumbo lutea [Willd.] Pers.). Unfortunately, the causal relationship between fly establishment and decline in hydrilla is frequently difficult to document. Declines may only be partial and localized. Detailed data are not available to document high levels of larvae in leaves of hydrilla before declines, and natural fluctuations in densities of submersed aquatic plants, such as hydrilla, are common. Economic Benefits Economic benefits of the introduced leaf-mining flies in the genus Hydrellia cannot yet be evaluated. The effects of these species are just now becoming visible and ongoing evaluation programs will be needed to measure any economic benefits procured. Recommendations for Future Work There are four major areas that should to be considered for future work: 1) domestic surveys to evaluate the current expansion and effect of the Hydrellia spp. flies that are already established; 2) assessing the influence abiotic and biotic factors have on establishment success and population build-up of these species; 3) developing improved methods for their mass rearing; and 4) conducting overseas surveys to locate previously identified and new biological control agents, especially in regions not studied previously. Continued field monitoring is needed to gain a clearer understanding of the potential impact of species of Hydrellia flies. This effort should include the development of lower cost, labor-efficient methods to measure hydrilla declines. Measuring changes in submersed plant populations has proven to be more difficult and costly than for terrestrial or floating plants. While range expansion of biological control agents is relatively easy to quantify, it is difficult to measure their impact since weed population changes occur over several growing seasons, with gradual replacement of hydrilla monocultures by mixtures of various native and non-native submersed plants (Grodowitz et al., 1999). Such evaluations are made even more difficult by the patchy distributions of these flies, which also can vary greatly between years at single locations. Reasons for such variation is unknown but could possibly be related to a complex of abiotic and biotic factors including overwintering conditions, plant nutritional variation, parasite loads, etc. For example, Grodowitz et al. (1995) cited that unusually cold weather and the lack of large releases was apparently the cause of declines in H. pakistanae populations in 1994 in Muscle Shoals, Alabama ponds.
costs per fly exceeding $0.50 per immature in greenhouse mass-rearing colonies (Freedman and Grodowitz, unpub.). Hence, a typical release of 50,000 individuals per site would cost more than $25,000 and be prohibitively expensive. Research to develop more cost effective rearing procedures is underway. For example, a mass-rearing facility based on the use of small ponds at an abandoned fish hatchery of the Tennessee Valley Authority Reservation in Muscle Shoals, Alabama, was highly successful (Grodowitz and Snoddy, 1995). A single harvest from a pond at this facility yielded more than 1.5 million flies and resulted in fly establishment throughout Lake Seminole, a large reservoir that borders both Florida and Georgia (Grodowitz, Cofrancesco, Stewart and Madsen, unpub.). While exact production costs are unknown it was significantly lower than the $0.50 per fly costs associated with greenhouse rearing techniques. Recently, a mass rearing system using a series of small ponds was implemented at the Lewisville Aquatic Ecosystem Research Facility in Lewisville, Texas. During 2000 and 2001 these ponds produced more than 600,000 individuals. Rearing costs were significantly lower, being less than $0.03 per immature (M. J. Grodowitz and R. Bare, unpub.). A similar, but smaller facility is currently under construction at the U.S. Army Engineer Research and Development Center in Vicksburg, Mississippi. Such facilities and procedures can significantly increase the number of sites at which releases can be made; however, local cooperation by state wildlife personnel and local water authorities is needed to facilitate the release of mass-reared flies.
Another area where more work is needed is in the understanding of the influence that abiotic and biotic factors have on fly establishment and population increase. Both laboratory and tank studies have quantified the influence of the plant’s nutritional composition on growth of Hydrellia spp. flies (Wheeler and Center, 1996; Doyle, Grodowitz, and Smart, unpub.). Tissue nutritional components can significantly affect fly survival, development times, fecundity, and female weight (an indicator of overall health). Nutritional components that appear to be important include nitrogen content and possibly phosphorus content, with higher levels increasing the overall health and vigor of the flies. Preliminary field data has indicated higher fly damage at sites with higher nitrogen levels (Wheeler and Center 2001; Grodowitz and Freedman, unpub.) but further information is needed to verify relationships between establishment success and population increase with plant nutritional composition. Among biotic factors of importance, more research is needed evaluating the impact of the pupal parasite Trichopria columbiana Ashmead, a diapriid wasp that attacks native Hydrellia species. Parasitism of the introduced Hydrellia species by T. columbiana can reach 30% by the end of the growing season in small ponds (Snell and Grodowitz, unpub.). However, the actual effect on fly population growth of removing 30% of the pupae from a given habitat is unknown. Also, T. columbiana may induce even higher mortality by probing pupae and hence causing mortality while searching for suitable oviposition sites (Bare and Grodowitz, unpub.). Highest priority for additional research needs to be given to the collection and study of new agents from overseas locations that attack permanently submersed hydrilla. Complexes of organisms that feed on and damage a variety of plant tissues are frequently needed to effectively suppress a target plant. In the case of hydrilla only one part of the plant, the leaves, are affected by established biological control agents. For efficient suppression, other agents are needed that, for example, could damage stems, roots, apical tips, turions, and/or tubers. Foreign exploration should target areas of the world that have received only limited previous attention, such as Southeast Asia. For example, several weevil species with potential as hydrilla herbivores have previously been identified (Table 2) but were never examined in any great detail. References Balciunas, J. K. 1982. Overseas searches for insects for control of aquatic plants, pp. 141-154. In Balciunas, J. K. 1983. Overseas searches for insects on hydrilla in Southeast Asia and Australia, pp. Balciunas, J. K. 1984. Overseas surveys of biocontrol agents for hydrilla, pp. 76-84. In Proceedings of Balciunas, J. K. 1985. Final report on the overseas surveys (1981-1983) for insects to control hydrilla. Balciunas, J. K. 1990. Biocontrol agents from temperate areas of Asia, pp. 25-33. In Miscellaneous Balciunas, J. K. and D. W. Burrows. 1996. Distribution, abundance, and field host-range of Hydrellia Balciunas, J. K. and T. D. Center. 1981. Preliminary host specificity tests of a Panamanian Balciunas, J. K. and P. P. Chen. 1993. Distribution of hydrilla in northern China: implications on future Balciunas, J. K. and D. H. Habeck. 1981. Recent range extension of hydrilla-damaging moth, Balciunas, J. K. and M. C. Minno. 1984. Quantitative survey of the insects and other macrofauna Balciunas, J. K. and M. C. Minno. 1985. Insects damaging hydrilla in the USA. Journal of Aquatic Balciunas, J. K. and M. F. Purcell. 1991. Distribution and biology of a new Bagous weevil Balciunas, J. K., T. D. Center, and F. A. Dray, Jr. 1989. Testing suitability of Australian bioagents for Balciunas, J. K., D. W. Burrows, and M. F. Purcell. 1996a. Australian surveys (1985-1992) for insect Balciunas, J. K., D. W. Burrows, and M. F. Purcell. 1996b. Comparison of the physiological and Baloch, G. M. and Sana-Ullah. 1974. Insects and other organisms associated with Hydrilla verticillata Baloch, G. M., A. G. Khan, and M. A. Ghani. 1972. Phenology, biology, and host-specificity of some Baloch, G. M., Sana-Ullah, and M. A. Ghani. 1980. Some promising insects for the biological control Bates, A. L. 1989. TVA aquatic plant research and cooperative efforts with the Aquatic Plant Control Bennett, C. A. 1986. My trip to Pakistan and India - weeds, weevils and worries. Aquatics 8: 9-11. Bennett, C. A. 1993. Quarantine biocontrol operations, pp. 88-92. In Miscellaneous Paper A-93-2. Bennett, C. A. and G. R. Buckingham. 1991. Laboratory biologies of Bagous affinis and B. laevigatus Bennett, C. A. and G. R. Buckingham. 1999. Biological control of hydrilla and Eurasian watermilfoil- Bennett, C. A. and G. R. Buckingham. 2000. The herbivorous insect fauna of a submersed weed, Bowes, G., A. S. Holaday, T. K. Van, and W. T. Haller. 1977. Photosynthetic and photorespiratory Buckingham, G. R. 1988. Reunion in Florida – hydrilla, a weevil, and a fly. Aquatics 10: 19-25. Buckingham, G. R. 1993. Foreign research on insect biocontrol agents, pp. 85-87. Proceedings of the Buckingham, G. R. 1994. Biological control of aquatic weeds, pp. 413-480. In Rosen, D., F. D. Buckingham, G. R. 1998. Surveys for insects that feed on Eurasian watermilfoil, Myriophyllum Buckingham, G. R. and J. K. Balciunas. 1994. Biological studies of Bagous hydrillae. Technical Report Buckingham, G. R. and C. A. Bennett. 1996. Laboratory biology of an immigrant asian moth, Buckingham, G. R. and C. A. Bennett. 1998. Host range studies with Bagous affinis (Coleoptera: Buckingham, G. R., E. A. Okrah, and M. C. Thomas. 1989. Laboratory host range tests with Hydrellia Buckingham, G. R., E. A. Okrah, and M. Christian-Meier. 1991. Laboratory biology and host range of CDFA (California Department of Food and Agriculture). 2000a. Hydrilla Program 20.20.01, Program CDFA (California Department of Food and Agriculture). 2000b. California Codes Food and Center, T. D. 1992. Release and field colonization of new biological control agents of Hydrilla Center, T. D., A. F. Cofrancesco, Jr., and J. K. Balciunas. 1990. Biological control of aquatic and Center, T. D., M. J. Grodowitz, A. F. Cofrancesco, G. Jubinsky, E. Snoddy, and J. E. Freedman. Chantaraprapha, N. and J. A. Litsinger. 1989. Host range and biology of three rice caseworms. Charudattan, R. 1973. Pathogenicity of fungi and bacteria from India to hydrilla and waterhyacinth. Charudattan, R. and D. E. McKinney. 1977. A Fusarium disease of the submersed aquatic weed, Charudattan, R., T. E. Freeman, R. E. Cullen, and F. M. Hofmeister. 1980. Evaluation of Fusarium Charudattan, R., T. E. Freeman, R. E. Cullen, and F. M. Hofmeister. 1984. Evaluation of Fusarium Cook, C. D. K. and R. Lüönd. 1982. A revision of the genus Hydrocharis (Hydrocharitaceae). Aquatic Cuda, J. P., B. R. Coon, Y. M. Dao, and T. D. Center. 2002. Biology and Laboratory Rearing of Cuda, J.P., B. R. Coon, J. L. Gillmore, and T. D. Center. 1999. Preliminary report on the biology of a Delfosse, E. S., B. D. Perkins, and K. K. Steward. 1976. A new U.S. record for Parapoynx diminutalis Deonier, D. L. 1971. A systematic and ecological study of nearctic Hydrellia (Diptera: Ephydridae). Dibble, E. D., K. L. Killgore, and S. L. Harrel. 1996. Assessment of fish-plant interactions. American Doyle, R. D., M. J. Grodowitz, R. M. Smart, and C. Owens. 2002. Impact of Herbivory by Hydrellia Drake, J. A., H. A. Mooney, F. di Castri, R. H. Groves, F. J. Kruger, M. Rejmanek, and M. Williamson Earhart, H. G. 1988. Highlights of the Potomac River and Chesapeake Bay, pp. 72-73. In Epler, J.H., J. P. Cuda, and T. D. Center. 2000. Redescription of Cricotopus lebetis (Diptera: Eubanks, M. J. 1987. South Atlantic Division, Mobile District, p. 11. In Miscellaneous Paper A-87-2. ERDC (U.S. Army Engineer Research and Development Center) 2001a. “Aquatic Plant Information ERDC (U.S. Army Engineer Research and Development Center) 2001b. “Noxious and Nuisance Plant FDEP (Florida Department of Evironmental Protection). 2000. Prohibited Aquatic Plants –HYPERLINK Farr, D. F., G. F. Bills, G. P. Chamuris, and A. Y. Rossman. 1989. Fungi on Plants and Plant Products Frankel, O. H., A. H. D. Brown, and J. J. Burdon. 1995. The Conservation of Plant Biodiversity. Godfrey, R. K. and J. W. Wooten. 1979. Aquatic Wetland Plants of Southeastern United States: Godfrey, K. E., L. W. Anderson, S. D. Perry, and N. Dechoretz. 1994. Overwintering and Godfrey, K. E., L. W. J. Anderson, C. E. Turner, K. Chang, D. Quimayousie, and J. Barajas. 1996. Grodowitz, M. J. and E. Snoddy. 1995. New pond facility for aquatic plant biocontrol research. Grodowitz, M. J., T. D. Center, and E. Snoddy. 1995. Current status of the use of insect biocontrol Grodowitz, M. J., T. D. Center, A. F. Cofrancesco, and J. E. Freedman. 1997. Release and Grodowitz, M. J., J. E. Freedman, A. F. Cofrancesco, and T. D. Center. 1999. Status of Hydrellia spp. Grodowitz, M. J., R. Doyle, and R. M. Smart. 2000a. Potential use of insect biocontrol agents for Grodowitz, M. J., J. E. Freedman, H. Jones, L. Jeffers, C. Lopez, and F. Nibling. 2000b. Status of Habeck, D. H. 1996. Australian moths for hydrilla control. Technical Report A-96-10. U.S. Army Haller, W. T. 1978. Hydrilla, a new and rapidly spreading aquatic weed problem. Agricultural Haller, W. T. 1982. Hydrilla goes to Washington. Aquatics 4: 6-7. Haller, W. T. and D. L. Sutton. 1975. Community structure and competition between Hydrilla and Harrel, S. L., E. D. Dibble, and K. J. Killgore. 2001. Foraging behavior of fishes in aquatic plants. Henderson, J. E. 1995. Use of economic information in the evaluation of aquatic plant control Joye, G. F. 1990. Biocontrol of Hydrilla verticillata with the endemic fungus Macrophomina phaseolina. Joye, G. F. and A. F. Cofrancesco, Jr. 1991. Studies on the use of fungal plant pathogens for control Joye, G. F. and R. N. Paul. 1992. Histology of infection of Hydrilla verticillata by Macrophomina Killgore, K. J., E. D. Dibble, and J. J. Hoover. 1993. Relationships between fish and aquatic plants: a Kirk, J. P., K. J. Killgore, J. V. Morrow, Jr., and J. W. Foltz. 1996. Triploid grass carp in Lake Marion, Kirk, J. P., J. V. Morrow, Jr., K. J. Killgore, S. J. de Kolowski, and J. W. Preacher. 2000. Population Krishnaswamy, S. and M. J. Chacko. 1990. Hydrellia spp. (Diptera: Ephydridae) attacking Hydrilla Langeland, K. A. 1990. Hydrilla (Hydrilla verticillata (L.f.) Royle): a continuing problem in Florida Langeland, K. A. and D. L. Sutton. 1980. Regrowth of hydrilla from axillary buds. Journal of Aquatic LDWF (Lousiana Department of Wildlife and Fisheries). 2000. Freshwater Recreational Fishing Madeira, P. T., T. K. Van, K. K. Steward, and R. J. Schnell. 1997. Random amplified polymorphic Madeira, P. T., C. C. Jacono, and T. K. Van. 2000. Monitoring hydrilla using two RAPD procedures Mahler, M. J. 1979. Hydrilla the number one problem. Aquatics 1: 56. Markham, R. H. 1986. Biological control agents of Hydrilla verticillata; final report on surveys in East Morrow, Jr., J. V., J. P. Kirk, and K. J. Killgore. 1997. Collection, age, growth, and population NCAWCA (North Carolina Aquatic Weed Control Act). 2000. North Carolina Plant Industry Division - O’Brien, C. and H. R. Pajni. 1989. Two Indian Bagous weevils (Coleoptera, Curculionidae), tuber OSDA (Oregon State Department of Agriculture). 2000. Oregon’s Quarantine Against Noxious Weeds. Pemberton, R. W. 1980. Exploration for natural enemies of Hydrilla verticillata in Eastern Africa. Pimm, S. L. 1991. The Balance of Nature?: Ecological Issues in the Conservation of Species and Rao, V. P. 1969. U.S. PL-480 Project: evaluation of natural enemies associated with witchweed, Sainty, G. R. and S. W. L. Jacobs. 1981. Waterplants of New South Wales. New South Wales Water SCDNR (South Carolina Department of Natural Resources). 2000. Aquatic Nuisance Species Schmitz, D. C. and D. Simberloff. 1997. Biological invasions: a growing threat. Issues in Science and Schmitz, D. C., B. V. Nelson, L. E. Nall, and J. D. Schardt. 1991. Exotic aquatic plants in Florida: a Shearer, J. F. 1996. Field and laboratory studies of the fungus Mycoleptodiscus terrestris as a Smart, R. M. and J. W. Barko. 1988. Effects of water chemistry on aquatic plants: interrelationships Speke, J. H. 1864. Journal of the Discovery of the Nile, 2nd ed. Blackwood and Sons, Edinborough, Steward, K. K., T. K. Van, C. Carter, and A. H. Pieterse. 1984. Hydrilla invades Washington, D. C., Tarver, D. P., J. A. Rodgers, M. J. Mahler, and R. L. Lazor. 1978. Aquatic Wetland Plants of Florida. TPWD (Texas Parks and Wildlife Department). 2000. Texas freshwater fishing: exotic fish, shellfish, USDA, NRCS (U.S. Department of Agriculture, Natural Resources Conservation Service). 1999. The Vandiver, Jr., V. V., T. K. Van, and K. K. Steward. 1982. Male hydrilla recently found in the United Varghese, G. and G. Singh. 1976. Progress in the search for natural enemies of hydrilla in Malaysia, Westman, W. E. 1990. Park management of exotic plant species: problems and issues. Conservation Wheeler, G. S. and T. D. Center. 1996. The influence of hydrilla leaf quality on larval growth and Wheeler, G. S. and T. D. Center. 2001. Impact of the biological control agent Hydrellia pakistanae WSDA (Washington State Department of Agriculture). 2000. Washington State Noxious Weed Laws, Zattau, W. C. 1988. Aquatic plant control operations support center, pp. 59-63. In Miscellaneous [ Contents ] [ Previous ] [ Next ] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ||||
| |
The Bugwood Network and Forestry Images Image Archive and Database Systems The University of Georgia - Warnell School of Forestry and Natural Resources and College of Agricultural and Environmental Sciences - Dept. of Entomology Last updated on Thursday, July 19, 2018 at 01:38 PM Questions and/or comments to the | |||