|
|
In: Van Driesche, R., et al., 2002, Biological Control of Invasive Plants in the Eastern United States, USDA Forest Service Publication FHTET-2002-04, 413 p. Pest Status of Weed Salvinia molesta D. S. Mitchell is a floating fern native to South America that in the last half of the twentieth century spread widely throughout the tropics and subtropics, moved in part by the trade in ornamental plants for fish tanks and ponds. It forms dense mats over lakes and slow moving rivers and causes large economic losses and a wide range of ecological problems to native species and communities. It is of interest in the United States because of its recent establishment in east Texas. Nature of Damage
waterways for transportation, cutting off access to important services, farm lands, and hunting grounds. The harm from salvinia mats to fisheries also can be very significant to communities dependent on fish for local consumption (sometimes as the main source of protein) or in areas where fish sales are the main source of cash income (Bennett, 1966; Thomas and Room, 1986). Salvinia also is a weed of paddy rice that reduces production by competing for water, nutrients and space (Anon., 1987).
by dense salvinia infestations. Water under mats of salvinia has a lower oxygen concentration (due to reduced surface area of water available for oxygenation, inhibition of photosynthesis by submerged plants, and consumption of dissolved oxygen by decaying salvinia), higher carbon dioxide and hydrogen sulphide concentrations, lower pH, and higher temperatures than nearby open water (Mitchell, 1978; Thomas and Room, 1986).
Through high growth rates and slow decomposition rates, salvinia reduces the concentration of nutrients that would otherwise be available to primary producers and organisms that depend on them (Sharma and Goel, 1986; Storrs and Julien, 1996). Mats of salvinia provide ideal habitat for Mansonia mosquitoes, a principal vector of rural elephantiasis in Sri Lanka (Pancho and Soerjani, 1978), and for other mosquito species involved in the transmission of encephalitis, dengue fever, and malaria (Creagh, 1991/92). Two species of Mansonia that occur in the United States, Mansonia dyari Belkin and Mansonia titillans (Walker), have been implicated in the transmission of St. Louis encephalitis and Venezuelan equine encephalitis, respectively (Lounibos et al., 1990). Extent of losses. The most detailed assessment of costs caused by salvinia was conducted in Sri Lanka using 1987 as the base year (Doeleman, 1989). Paddy rice losses, fishing losses, other losses (power generation, transport, washing and bathing, etc.), health costs, abatement costs, and economic benefits were considered. No environmental costs were included, but they were recognized as important. There were no identified benefits from salvinia. Total costs associated with salvinia were estimated to be between 24.7 million and 56.7 million rupees (in Australian dollars, between 0.9 and 2.1 million) for 1987. This information was used to determine the benefits from biological control over the following 25 years. The benefits were 53 rupees or dollars per rupee or dollar invested, or 1,673 man-hours per man-hour invested. Using this information as a guide, Room and Julien (1995) estimated that the annual benefits gained from successful biological control of salvinia worldwide were approximately $150 million U.S. Geographical Distribution The native range of salvinia is an area in southeastern Brazil (Forno and Harley, 1979). Its first recorded exotic establishment was in Sri Lanka in 1939 (Williams, 1956). It has since become established in India (Cook and Gut, 1971), Australia (Room and Julien, 1995), Papua New Guinea (Mitchell, 1979), Cuba, Trinidad, Guyana, Columbia (Holm et al., 1979), South Africa (Cilliers, 1991), Botswana (Edwards and Thomas, 1977), Kenya, Zambia (Mitchell and Tur, 1975), Namibia (Forno and Smith, 1999), Madagascar (Room and Julien, 1995), Ghana and Cote D’Ivoire (M. Julien, pers. obs.), Indonesia (Java, Borneo, Sulawesi), Malaysia (mainland Sabah, Sarawak) (R. Chan, pers. comm.), the Philippines (Pablico et al., 1989), Fiji (Kamath, 1979), and New Zealand (Randall, 1996). Salvinia was first reported outside of cultivation in the United States in 1995 at a pond in southeastern South Carolina (Johnson, 1995). It was eradicated before spread occurred. It was next found in Houston, Texas, in May 1998, and then at other sites in Texas and in Louisiana during 1998. During 1999 it was found in ponds and rivers in Alabama, Arizona, California, Florida, Georgia, Hawaii, Mississippi, and Oklahoma (Jacono et al., 2000; see also Jacono’s web site). Salvinia is readily available for purchase in the United States, particularly through the Internet. Background Information on Pest Plant Taxonomy The aquatic fern family Salviniaceae is placed within the order Hydropteridales and consists of a single genus, Salvinia. Ten species of Salvinia occur worldwide (Herzog, 1935; de la Sota, 1962, 1963, 1964, 1982; Mitchell, 1972). None are native to the United States (Jacono et al., 2000) although seven species
Growth and reproduction. Salvinia is pentaploid, has a chromosome number of 45, and is incapable of sexual reproduction (Loyal and Grewal, 1966). Each node bears a series of up to three axillary buds that develop successively under normal growing conditions (Room, 1988), and up to six in response to damage (Julien and Bourne, 1986). The number of axillary buds that grow, the rate of growth, and plant size are largely dependent on available nutrients. Growth is apically dominant and progresses by expansion of apical and axillary buds, the latter forming branches. New plants form when older plants break apart due to senescence or damage (Room, 1983). Factors affecting growth. Salvinia is a perennial plant with no seasonal periodicity, although changes in growth may be related to seasonal variations such as changes in temperature. Salvinia is well adapted to growth in low nutrient waters and can take up nutrients quickly when they become available (Room and Thomas, 1986). The proportion of axillary buds that develop is correlated with the nitrogen content of the plant (Room, 1983; Julien and Bourne, 1986), and the nitrogen content increases following removal of buds by insects or other agents (Room and Thomas, 1985; Julien and Bourne, 1986; Forno and Semple, 1987). At low levels of nitrogen leaves are larger, ‘roots’ longer, sporocarps occur more frequently, and rhizome branching is reduced (Room, 1983; Julien and Bourne, 1986; Room, 1988; Room and Julien, 1995). The nitrogen content of salvinia ranges from 0.6 to 4.0% dry weight (Room and Thomas, 1986). The maximum rate of nitrogen uptake, calculated from rates of growth, is near 8 mg nitrogen/g dry weight of salvinia/day or about 6,000 kg nitrogen/ha/year (Room, 1986). Actual measurements at a sewage treatment lagoon indicated an uptake of 1,580 kg nitrogen/ha/year (Finlayson et al., 1982). The optimum temperature for growth is 30°C. Room (1986) described the effect of temperature, above and below 30°C, on relative growth rates and predicted no growth below 10°C and above 40°C. Temperature does not affect the proportion of axillary buds that expand to initiate new branches (Room, 1988). Exposure to temperatures below –3°C or above 43°C for more than two hours kills salvinia (Whiteman and Room, 1991). Leaf temperatures that exceeded 40°C and sometime approached 50°C for the hottest parts of days did not obviously affect growth, but water temperatures remained below 40°C and probably acted as a heat sink for the plants (Storrs and Julien, 1996). Plants may be killed by frost but protected parts and unfrozen buds survive. Water bodies are normally cooler than the air in summer and warmer in winter due to thermal inertia. This helps protect salvinia from temperature extremes. Mats of salvinia can grow in water bodies with conductivities ranging from 100 µS/cm to 1,400 µS/cm (Mitchell et al., 1980; Room and Gill, 1985). In water with 10% of the salinity of seawater (4,800 µS/cm), growth was reduced by 25% (Divakaran et al, 1980); at 20% salinity, growth was very slow; while at 30%, plants died after 30 minutes exposure (Room and Julien, 1995). Optimum pH for growth is 6.0 (Cary and Weerts, 1984). In the field the plant grows at pH values from 5.2 to 9.5 (Holm et al., 1977; Mitchell et al., 1980). Salvinia compensates for the destruction of buds by initiating growth of dormant buds. Complete compensation occurs only when high levels of nitrogen are available (Julien and Bourne, 1986; Julien et al., 1987). Destruction of leaves (Julien and Bourne, 1988) and rhizomes (Julien and Bourne, 1986) does not induce compensatory growth. Growth rates and density. On Lake Kariba, Zimbabwe, numbers of leaves (ramets) doubled in eight to 17 days (Gaudet, 1973; Mitchell and Tur, 1975). In the Kakadu National Park, Australia, dry weight doubled in five to 30 days (Storrs and Julien, 1996). Under ideal growth conditions, biomass and numbers of ramets typically double in two to three days (Mitchell and Tur, 1975; Cary and Weerts, 1983). Densities from as high as 2,500 large tertiary form ramets per m2 (in nutrient-poor water) to 30,000 small tertiary form ramets (in nutrient-rich waters) have been noted. At these densities, natality is equaled by mortality (Room and Julien, 1995). Salvinia is 95% water by weight and biomass of living shoots can exceed 600 g/m2 of dry weight, while biomass of living and dead shoots and ‘roots’ may exceed 1,600 g/m2 of dry weight or 400 t/ha of fresh weight (Room and Julien, 1995). Fresh weight biomass in Texas varied through the year, reaching a high in October 1999 of 248 t/ha and a low of 84 t/ha in January 2000 (P. Tipping, unpub. data). Spread. Salvinia is spread within and between aquatic systems mainly by people. It is spread accidentally when equipment or boats are moved and deliberately when it is used as a pond, aquarium, or water-garden plant or as a biological weapon (Gewertz, 1983). It is carried on animals as they move from infested water bodies (Forno and Smith, 1999). Dispersal within a water body or catchment is by wind and water currents (Room and Julien, 1995). Currents and floods wash mats away and growth is best in still or slow moving water. In its native range in southeastern Brazil, salvinia is a component of floating and emergent plant communities. Salvinia supports a variety of natural enemies (Forno and Bourne, 1984), and it normally does not form the extensive mats prevalent in its exotic range. Analysis of Related Native Plants in the Eastern United States The Salviniaceae are included within a monophyletic clade of heterosporous genera that also encompasses the Azollaceae and Marsileaceae (Pryer and Smith, 1998; Pryer et al., 1995; Hasebe et al., 1995), all aquatic leptosporangiate ferns. This clade includes ten species in the North American flora. The Azollaceae once were included within the Salviniaceae, but the relationship is not close and they have since been separated (Lumpkin, 1993). Azollaceae consists of the single genus Azolla. There are three species of Azolla – Azolla caroliniana Willdenow, Azolla mexicana C. Presl, and Azolla filiculoides Lamarck – that are native to North America (Lumpkin, 1993). The Marsileaceae includes two genera, Marsilea and Pilularia, both of which occur in North America. Seven species within these two genera are part of the North American flora (Johnson, 1993): Marsilea quadrifolia Linnaeus, Marsilea ancylopoda A. Braun, Marsilea oligospora Gooding, Marsilea mollis B. L. Robinson and Fernald, Marsilea macropoda Engelmann, Marsilea vestita Hooker and Greville, and Pilularia americana A. Braun. Marsilea quadrifolia, a native of Europe and Asia, is introduced and M. ancylopoda is extinct, so only eight native species remain.
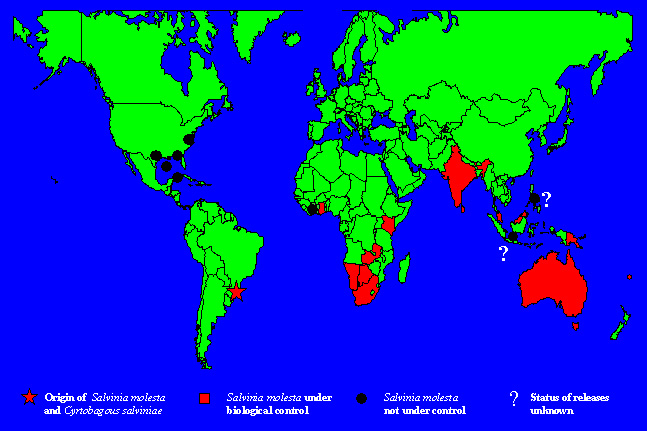
Areas Surveyed for Natural Enemies The first surveys for potential biological control agents for S. molesta were conducted in Trinidad, Guyana and northeastern Brazil from 1961 to 1963 (Bennett, 1966), and in Argentina prior to 1975 (Bennett, 1975), where species in the S. auriculata complex other than S. molesta occur. At this time the true identity and the native range of S. molesta were not known. Surveys for natural enemies were conducted in Trinidad, Venezuela, Guyana, Uruguay, Paraguay, Brazil, and Argentina during 1978 to 1981. In 1978, the previously unknown range of S. molesta was identified (Forno and Harley, 1979), permitting surveys to focus on the relatively small native range of the target weed rather than the larger range of the S. auriculata complex (Forno and Bourne, 1984). Natural Enemies Found The natural enemies of S. molesta and the related species in the S. auriculata complex are listed in Forno and Bourne (1984), including species collected by Bennett (1975). Twenty-five phytophagous or possibly phytophagous species have been recorded from S. molesta, compared to 49 species from the four species of the S. auriculata complex. Four of these species have been used as biological control agents against S. molesta. The first three, Cyrtobagous singularis Hustache, Paulinia acuminata (De Geer), and Samea multiplicalis (Guenée) (identified during the early exploration [Bennett, 1966]) have not been successful control agents. The fourth, Cyrtobagous salviniae Calder and Sands, was found during later work (Sands, 1983) and has been extremely successful. Host Range Tests and Results Host range studies on the three unsuccessful species are summarized in Bennett (1966), Sankaran and Ramaseshiah (1973), Sands and Kassulke (1984, 1986), and Knopf and Habeck (1976). Host range tests to assess feeding by C. salviniae were carried out in Australia on 46 species from six families of Pteridophyta (ferns), eleven families of Monocotyledons, and sixteen families of Dicotyledons (Forno et al., 1983). (This weevil was later found to be a new species and subsequently descrbed as C. salviniae Calder and Sands). Test plants were exposed to mature weevils in three replicates in choice tests. Adult feeding occurred on Pistia stratiotes L., but the insect was unable to reproduce on that species. Minor leaf feeding was observed in choice tests on Ipomea batatas (L.) Lam. (sweet potato) when the leaves were held in contact with water, an abnormal condition. Adults failed to feed on I. batatas in no-choice tests in a non-aquatic situation and died within seven days. Host specificity tests indicated that this weevil was restricted to S. molesta. It has never been observed attacking plants other than Salvinia species in the field in South America, including those that grew in association with S. molesta such as water fern (Azolla sp.), waterhyacinth (Eichhornia crassipes [Mart.] Solms-Laubach), and waterlettuce (P. stratiotes) (Forno et al., 1983). Importantly, this weevil has not been found to attack any other plants even when huge populations were starving following population crashes of salvinia. Releases Made The grasshopper P. acuminata, collected from S. auriculata in Trinidad, was released in Zimbabwe (in 1969 and again in 1971), Kenya and Zambia (1970), Botswana (1971 and 1975), Sri Lanka (1973 and 1978), India (1994), and Fiji (1975). It failed to establish in Botswana, Kenya, and Sri Lanka and does not provide control in the countries where it established. The weevil C. singularis, collected from S. auriculata in Trinidad, was released in Botswana (in 1971 and 1976), Zambia (1971), and Fiji (1976). It is established in each country but is not providing control. The moth S. multiplicalis, also collected from S. auriculata in Trinidad, was released in Zambia (1970), Botswana (1972), and Fiji (1976). It failed to establish in Fiji and Zambia and does not control the weed in Botswana (Julien and Griffiths, 1998). It was later collected from S. molesta in Brazil and released in Australia during 1981, where it established widely but failed to provide control (Room et al., 1984; Forno, 1987). Releases of C. salviniae from S. molesta in southeastern Brazil were made first in Australia in 1980. This weevil now controls the weed in most tropical and subtropical areas (Fig. 5) and in some temperate climates (Fig. 6). It has been released in 15 countries and controls the weed in at least 12 of these (Table 1). In Cote D’Ivoire, insufficient time has elapsed to measure success. Information on project results is not available for the Philippines or Indonesia. Map 1 shows the countries that have or have had serious salvinia problems and those where biological control has been successful.
Table 1. The Status of Releases of Cyrtobagous Salviniae Calder and Sands for Each Country and the Date of Initial Release (Modified from Julien and Griffiths, 1998)
History of Biological Control Efforts in the Eastern United States A weevil found attacking S. minima in Florida was identified as C. singularis (Kissinger, 1966), but Calder and Sands (1985) listed Cyrtobagous specimens from Florida as C. salviniae and did not consider the range of C. singularis to include North America. When S. molesta was found in the United States and biological control was considered, further morphological examination of weevils from Florida suggested that they were C. salviniae (C. O’Brien, pers. comm.). This weevil was collected from S. minima in Florida by scientists from the USDA, ARS Invasive Plant Research Laboratory and released at sites in Liberty, Bridge City, and Toledo Bend Reservoir in Texas, and at Salter Creek in Louisiana during 1999. When another USDA laboratory planned to release an Australian population of C. salviniae, a molecular comparison of the D2 gene was made between Florida and Australian material to provide a means of distinguishing between weevils from the two sources. Several base pair differences were found but the taxonomic significance of this is not yet clear. Further studies utilizing molecular, morphological and bioassay methods are planned. Until these differences are explained, further release of the Florida population will be suspended. No release of Australian material has been done yet. Biology and Ecology of Key Natural Enemies Salvinia weevils, Cyrtobagous salviniae and C. singularis (Coleoptera: Curculionidae)
explained why one species failed to cause significant damage to the weed while the other proved to be an excellent control agent. Larvae of C. salviniae tunnel within the rhizomes causing them to disintegrate. Larvae also tunnel in the buds and adults eat buds, thus suppressing growth and vegetative propagation of this sterile weed. Larvae and adults of C. singularis feed on leaves and other tissues but don’t affect the rhizomes or meristems. This research provided a classic example of the importance of careful taxonomic study of both the weed and the insects for successful biological control (Thomas and Room, 1986). Other important differences between the two species are that C. salviniae has a higher intrinsic rate of increase, lays seven times more eggs, and oviposits with greater frequency. Furthermore, this species’ oviposition is less affected by changes in the nutrient status of the host plant, and larval and pupal survival rates are higher (Sands et al., 1986).
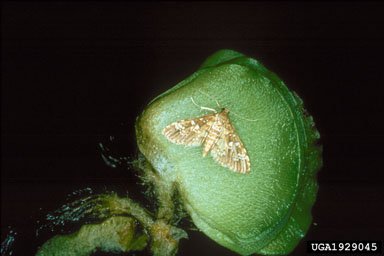
C. salviniae. The adult male of C. salviniae (1.8 x 0.9 mm) is slightly smaller than the female (2.2 x 1.2 mm). Newly emerged adults are brown, darkening to black in about five days. Detailed descriptions are given in Calder and Sands (1985) of the features that distinguish this species from C. singularis. Adults are found on or beneath young leaves, on or inside the developing leaves or among ‘roots’. When under water, adults respire by means of an air bubble (called a plastron) that adheres to their ventral surface (Forno et al., 1983). Multiple matings occur five to 26 days after emergence. At 25.5ºC, oviposition begins after six to 14 days. Eggs (0.5 x 0.24 mm) are laid singly in cavities excavated by adults in lower leaves, developing leaves, rhizomes, and ‘roots.’ At 25.5ºC, females lay one egg every two to five days for at least 60 days (Forno et al.., 1983). Eggs hatch in 10 days at this temperature. Newly emerged larvae (1 mm) are white. They feed initially on ‘roots’ in or on the small buds, and later inside rhizomes, completing three instars in approximately 23 days (Forno et al., 1983). Development rate is dependent on temperature and the nutrient status of the host plant, larval development taking 13.4 days at 31ºC on ‘high’ nitrogen plants. Larvae prefer to tunnel in young rhizomes and more tunneling occurs if plants are low in nitrogen. Larvae do not survive below 16.3°C (Sands et al., 1983). Pupation occurs in a cocoon (2 x 2.6 mm), which is woven from ‘root hairs’ and attached underwater to the ‘roots,’ rhizomes or leaf bases. At 25.5ºC, pupae require 12.6 days for full development. Pupal duration is not affected by plant quality (Forno et al., 1983; Sands et al., 1983). Oviposition does not occur below 21ºC,and eggs fail to hatch below 20ºC or above 36ºC. Adults feed between 13ºC and 33ºC (Forno et al., 1983). The lower lethal temperature at which 50% of the adult population would be expected to die is -5.2ºC (Reaney 1999). Population densities of C. salviniae are capable of reaching 300 adults and 900 larvae per m2, levels estimated as necessary for control (Room, 1988, 1990; Room and Thomas, 1985). Weevils in the genus Cyrtobagous were first recorded from the United States in Florida at the Archbold Biological Station (Highlands County) in 1962 (Kissinger, 1966). It is assumed that these weevils were accidentally introduced from South America, because of the lack of any earlier U.S. records and the adventive status of its host plant, S. minima. Kissinger (1966) considered the Florida weevils to be C. singularis, but this was before C. salviniae was recognized as a separate species. Calder and Sands (1985) later classified the Florida specimens as C. salviniae, but noted that the C. salviniae from S. minima in Florida were significantly smaller than those from S. molesta in Brazil. Based solely on morphological features, the weevils from Florida (later released in Texas) seem to be C. salviniae (C. O’Brien, pers. comm.). However, recent DNA assessments suggest that that the Florida material differs from C. salviniae from Australia in some respects (Goolsby et al., 2000). Whether these differences imply separate species status is under investigation. C. singularis. The biology of C. singularis is less well known and it is presumably similar to C. salviniae. For morphological differences between adults of this species and those of C. salviniae see Calder and Sands (1985), and for larvae, May and Sands (1986). Feeding differences between the species are outlined in Sands and Schotz (1985), and other differences in life history and intrinsic rates of increase are discussed in Sands et al. (1986). Adults are small (2 to 3 mm), black, sub-aquatic weevils that reside on or beneath leaves. While under water they respire using a plastron. Adults preferentially feed on apical leaves but also on the second to fifth pair of leaves, buds, and petioles (Sands and Schotz, 1985). Eggs are laid singly in cavities made by females in leaves. Unlike C. salviniae, whose larvae feed internally, those of C. singularis feed on the outer surfaces of submerged buds, rhizomes, and petioles. Feeding results in bud destruction, but not rhizome disintegration, and plants retain their capacity for regrowth (Sands and Schotz, 1985). Field population densities of C. singularis do not exceed 50 adults per m2 (Schlettwein, 1985), a level that is insufficient to significantly damage salvinia (Room, 1990). This insect has not been a useful biological control agent (Julien and Griffiths, 1998). Waterlettuce moth, Samea multiplicalis (Lepidoptera: Pyralidae) Larvae and adults of S. multiplicalis are very similar to the closely related species, Niphograpta (Sameodes) albiguttalis (Warren) (waterhyacinth moth, Pyralidae). Center et al. (1982) give characters to separate larvae of these species. Samea multiplicalis was originally described from Brazil where it was observed feeding on waterhyacinth. It is widely distributed throughout warmer regions of North and South America. In Florida, it is most commonly found on waterlettuce but also is present on A. caroliniana and S. minima. It is occasionally abundant on small waterhyacinth plants, feeding within inflated leaf petioles (Knopf and Habeck, 1976). This species was introduced into Australia for biological control of both S. molesta and P. stratiotes (Sands and Kassulke, 1984). Adults (Fig. 8) are 6.5 to 10.5 mm long (Sands and Kassulke, 1984), tan, with brown and cream markings on both fore and hind wings. Females lay approximately 300 eggs during their four-to-seven-day life span (Knopf and Habeck, 1976; Sands and Kassulke, 1984; Taylor, 1984). Moths prefer to lay eggs on undamaged salvinia plants with high nitrogen content (Taylor and Forno, 1987). Most eggs are laid singly among the epidermal plant hairs on the lower surfaces of waterlettuce leaves, on the upper surfaces of Salvinia leaves, or lodged between the leaves of Azolla species. Eggs hatch in about four days at 26ºC. Larvae (Fig. 9) may feed from within a refugium (made of silk and plant hair) attached to the external leaf surface or, for waterlettuce, within galleries in the leaves. Larvae periodically extend the refugium to reach fresh leaf material (Knopf and Habeck, 1976). Larger larvae feed on the buds of plants, often killing the growing apex. Larvae also will eat mature waterlettuce fruits and consequently destroy enclosed seeds.
On salvinia, temperature and plant quality interact to determine rates of insect growth, number of larval instars, fecundity, and survival (Taylor and Sands, 1986; Taylor, 1984, 1988, 1989). Optimum temperature for development for all stages is 28 to 30ºC. Fecundity is greatest at 20 to 22ºC and egg survival highest at 25 to 26ºC (Taylor, 1988). Larvae complete development in 17 to 35 days on salvinia (Sands and Kassulke, 1984; Taylor, 1984). Development on waterlettuce and other salvinia species is described in Bennett (1966) and Knopf and Habeck (1976). Pupation occurs within a silken cocoon. On waterlettuce, this cocoon is usually formed within the spongy portion of a leaf but on S. molesta it is constructed among old leaves. Pupal development requires four to seven days at 28ºC on waterlettuce and S. minima (Knopf and Habeck, 1976) and eight to nine days at 26ºC on S. molesta (Sands and Kassulke, 1984). Total development (egg to adult) requires 24.6 (Knopf and Habeck, 1976) to 42 days (Sands and Kassulke, 1984). Intrinsic rates of increase are highest in autumn and decline in summer and winter (Room et al., 1984; Taylor, 1988). Levels of parasitism (24%) and disease in Australian populations on salvinia do not explain the seasonal variation in population growth rates (Semple and Forno, 1987); rather, field population densities are strongly determined by temperature and the nutritional quality of the plant (Taylor, 1988). Salvinia grasshopper, Paulinia acuminata (Orthoptera: Pauliniidae)
are lower (Thomas, 1980). Eggs take 17 to 20 days to hatch; six nymphal instars complete development after 47 days; the pre-ovipositional period takes eight to 10 days; and duration from egg to adult is 67 days on average (Sands and Kassulke, 1986).
Females live 50 days or more and lay 200 or more eggs. Adults usually are brachypterous but macropterous forms occasionally occur in the field and are frequent in laboratory cultures (Bennett, 1966). Adults and nymphs feed on all plant parts above the water level when food is scarce. Grasshoppers prefer to feed on new growth and oviposition is reduced when the weed is matted (Mitchell and Rose, 1979). Under extremely hot conditions adults shelter in the water with their heads exposed (Thomas, 1980). In the field at Lake Kariba, where the mean annual temperature was 24 to 28ºC, it was estimated that P. acuminata could complete three generations per year (Thomas, 1980). A significant reduction in salvinia on Lake Kariba occurred in 1972/3, following the release of P. acuminata in 1970. However, the decline of the weed at this location has been attributed to other factors such as nutrient stress on the weed (Mitchell and Rose, 1979; Marshall and Junor, 1981). Chisholm (1979) demonstrated that densities of more than 85 P. acuminata per m2 feeding for 24 days were required to reduce production of new leaves. On Lake Kariba during the period when salvinia was a problem the maximum grasshopper density recorded was 27 per square meter (Marshall and Junor, 1981), suggesting that the insect might not have been a primary cause of decline in the weed. Sands and Kassulke (1986) noted that field populations in Africa of 45 to 54 P. acuminata per m2 grazed up to 87% of leaves without affecting apical or lateral growth or killing plants. Evaluation of Project Outcomes In the United States, S. molesta is a new problem so evaluations are premature. As mentioned previously, C. salviniae, a naturalized species on S. minima in Florida, was released in Texas in 1999. It will take another year to determine if establishment has occurred. Recommendations for Future Work Currently, three release sites and three insect-free control sites are being monitored monthly in water bodies in eastern Texas and western Louisiana infested with S. molesta using standard protocols. In addition, molecular techniques are being utilized to identify and compare at least six salvinia species, including those outside of the S. auriculata complex. This will provide a means of identifying Salvinia species should any new invasions occur in the United States or elsewhere. Similar molecular testing of Cyrtobagous sp. collected from different salvinia species may provide insights to the taxonomy of this group and their associated host ranges. We anticipate that the same favorable results as obtained in at least 12 countries on three continents can be repeated in the United States, namely, reductions in the density and abundance of S. molesta, to acceptable levels. Reductions may range from local extinctions to maintenance of small populations of the weed along the fringes of ponds, lakes, and rivers. Acknowledgements We thank Drs. Tim Heard and Peter Room for reviewing the manuscript and Dr. Peter Room for preparing the world map. We also thank Dr. Peter Room and Mr. Richard Chan for permission to use their photographs. References Anonymous. 1987. Indonesia girds to battle Salvinia molesta. Weedwatcher 2: 1-2. (Publication of Bennett, F. D. 1966. Investigations on the insects attacking the aquatic ferns, Salvinia spp. in Trinidad Bennett, F. D. 1975. Insects and plant pathogens for the control of Salvinia and Pistia, pp. 28-35. In Bonnet, A. L. M. 1955. Contribution a l’etude des hydropteridees: recherches sur Salvinia auriculata Calder, A. A. and D. P. A. Sands. 1985. A new Brazilian Cyrtobagous Hustache (Coleoptera: Cary, P. R. and P. G. J. Weerts. 1983. Growth of Salvinia molesta as affected by water temperature Cary, P. R. and P. G. J. Weerts. 1984. Growth of Salvinia molesta as affected by water temperature Center, T. D., J. K. Balciunas, and D. H. Habeck. 1982. Descriptions of Sameodes albiguttalis Chisholm, I. F. 1979. A laboratory investigation of Paulinia acuminata (De Geer) (Orthoptera: Cilliers, C. J. 1991. Biological control of water fern, Salvinia molesta (Salviniaceae), in South Africa. Cook, C. D. K. and B. J. Gut. 1971. Salvinia in the State of Kerala, India. Pest Articles and News Creagh, C. 1991/92. A marauding weed in check. Ecos 70: 26-29. Croxdale, J. G. 1978. Salvinia leaves. I. Origin and early differentiation of floating and submerged Croxdale, J. G. 1979. Salvinia leaves. II. Morphogenesis of the floating leaf. Canadian Journal of Croxdale, J. G. 1981. Salvinia leaves. III. Morphogenesis of the submerged leaf. Canadian Journal of de la Sota, E. R. 1962. Contribución al conocimiento de las Salviniaceae neotropicales. I. Salvinia de la Sota, E. R. 1963. Contribución al conocimiento de las Salviniaceae Neotropicales. IV. Data de la Sota, E. R. 1964. Contribución al conocimiento de las Salviniaceae neotropicales. V. Salvinia de la Sota, E. R. 1976. Sinopsis de las especies Argentinas del genero Salvinia Adanson de la Sota, E. R. 1982. Sobre Salvinia radula Baker (Salviniaceae-Pteridophyta) Biotica 7: 457-462. Divakaran, O., M. Arunachalam, and N. B. Nair. 1980. Growth rates of Salvinia molesta Mitchell with Doeleman, J. A. 1989. Biological Control of Salvinia molesta in Sri Lanka: An Assessment of Costs Edwards, D. and P. A. Thomas. 1977. The Salvinia molesta problem in the northern Botswana and Finlayson, C. M., T. P. Farrell, and D. J. Griffiths. 1982. Treatment of Sewage Effluent Using the Forno, I. W. 1983. Native distribution of the Salvinia auriculata complex and keys to species Forno, I. W. 1987. Biological control of the floating fern Salvinia molesta in north-eastern Australia: Forno, I. W. and A. S. Bourne. 1984. Studies in South America of arthropods on the Salvinia Forno, I. W. and K. L. S. Harley. 1979. The occurrence of Salvinia molesta in Brazil. Aquatic Botany Forno, I. W. and J. L. Semple. 1987. Response of Salvinia molesta to insect damage: changes in Forno, I. W. and P. A. Smith. 1999. Management of the alien weed, Salvinia molesta, in the wetlands Forno, I. W., D. P. A. Sands, and W. Sexton. 1983. Distribution, biology and host specificity of Gaudet, J. J. 1973. Growth of a floating aquatic weed, Salvinia, under standard conditions. Gewertz, D. B. 1983. Salvinia molesta: the destruction of an ecosystem, pp. 196-217. In Gewertz, Goolsby, J. A., P. W. Tipping, T. D. Center, and F. Driver. 2000. Evidence of a new Hasebe, M., P. G. Wolf, K. M. Pryer, K. Ueda, M. Ito, R. Sano, G. J. Gastony, J. Yokoyama, Herzog, T. K. J. 1935. Ein beitrag zur systematik der gattung Salvinia. Hedwigia 74: 257-284. Holm, L. G., D. L. Plucknett, J. V. Pancho, and J. P. Herberger. 1977. The World’s Worst Weeds. Holm, L. G., J. V. Pancho, J. P. Herberger, and D. L. Plucknett. 1979. A Geographical Atlas of World Jacono, C. C., T. R. Davern, and T. D. Center. 2000. The adventive status of Salvinia minima and S. Jacono, C. C. Salvinia molesta, giant salvinia. http://salvinia.er.usgs.gov, (accessed March 26, 2002). Johnson, D. M. 1993. Marsileaceae Mirbel – Water clover family, pp. 331-335. In Flora of North Johnson, D. 1995. Giant salvinia found in South Carolina. Aquatics 17: 22. Julien, M. H. and A. S. Bourne. 1986. Compensatory branching and changes in nitrogen content in Julien, M. H. and A. S. Bourne. 1988. Effects of leaf-feeding by larvae of the moth Samea multiplicalis Julien, M. H. and M. W. Griffiths (eds.). 1998. Biological Control of Weeds. A World Catalogue of Julien, M. H., A. S. Bourne, and R. R. Chan. 1987. Effects of adult and larval Cyrtobagous salviniae Kamath, M. K. 1979. A review of biological control of insect pests and noxious weeds in Fiji (1969- Kissinger, D. G. 1966. Cyrtobagous Hustache, a genus of weevils new to the United States fauna Knopf, K. W. and D. H. Habeck. 1976. Life history and biology of Samea multiplicalis. Environmental Lounibos, L. P., V. L. Larson, and C. D. Morris. 1990. Parity, fecundity, and body size of Mansonia Loyal, D. S. and R. K. Grewal. 1966. Cytological study on sterility in Salvinia auriculata Aublet with a Lumpkin, T. A. 1993. Azollaceae Wettstein – Azolla family, pp. 338-342. In Flora of North America May, B. M. and D. P. A. Sands. 1986. Descriptions of larvae and biology of Cyrtobagous (Coleoptera: Mitchell, D. S. 1972. The Kariba weed: Salvinia molesta. British Fern Gazette 10: 251-252. Mitchell, D. S. 1978. Aquatic Weeds in Australian Inland Waters. Australian Government Publishing Mitchell, D. S. 1978/9. Aquatic weeds in Papua New Guinea. Science in New Guinea 6: 154-160. Mitchell, D. S. 1979. The incidence and management of Salvinia molesta in Papua New Guinea. Draft Mitchell, D. S. and D. J. W. Rose. 1979. Factors affecting fluctuations in extent of Salvinia molesta on Mitchell, D. S. and P. A. Thomas. 1972. The Taxonomy and Distribution of Salvinia Species in the Mitchell, D. S. and N. M. Tur. 1975. The rate of growth of Salvinia molesta [S. auriculata Auct.] in Mitchell, D. S., T. Petr, and A. B. Viner. 1980. The water fern Salvinia molesta in the Sepik River, Pablico, P. P., L. E. Estorninos Jr., E. M. Castin, and K. Moody. 1989. The occurrence and spread of Pancho, J. V. and M. Soerjani. 1978. Aquatic Weeds of Southeast Asia. National Publication Pryer, K. M. and A. R. Smith. 1998. Leptosporangiate ferns. In Madison, D. R. and W. P. Madison Pryer, K. M., A. R. Smith, and J. E. Skog. 1995. Phylogenetic relationships of extant ferns based on Randall, J. L. 1996. National pest management strategy for noxious weeds. The Royal Society of Reaney, L. 1999. The suitability of Cyrtobagous salviniae as a biological control agent for the aquatic Room, P. M. 1983. Falling-apart as a lifestyle – the rhizome architecture and population growth of Room, P. M. 1986. Equations relating growth and uptake of nitrogen by Salvinia molesta to Room, P. M. 1988. Effects of temperature, nutrients and a beetle on branch architecture of the Room, P. M. 1990. Ecology of a simple plant-herbivore system: biological control of salvinia. Trends Room, P. M. and J. Y. Gill. 1985. The chemical environment of Salvinia molesta Mitchell: ionic Room, P. M. and M. H. Julien. 1995. Salvinia molesta D.S. Mitchell, pp. 217-230. In Groves, R. H., Room, P. M. and P. A. Thomas. 1985. Nitrogen and establishment of a beetle for biological control of Room, P. M. and P. A. Thomas. 1986. Nitrogen, phosphorus and potassium in Salvinia molesta Room, P. M., I. W. Forno, and D. P. A. Sands. 1984. Establishment in Australia of two insects for Sands, D. P. A. 1983. Identity of Cyrtobagous sp. (Coleoptera: Curculionidae) introduced into Sands, D. P. A. and R. C. Kassulke. 1984. Samea multiplicalis (Lep.:Pyralidae), for biological control Sands, D. P. A. and R. C. Kassulke. 1986. Assessment of Paulinia acuminata (de Geer) (Orthoptera: Sands, D. P. A. and M. Schotz. 1985. Control or no control: A comparison of the feeding strategies of Sands, D. P. A., M. Schotz, and A. S. Bourne. 1983. The feeding characteristics and development of Sands, D. P. A., M. Schotz, and A. S. Bourne. 1986. A comparative study on the intrinsic rates of Sankaran, T. and G. Ramaseshiah. 1973. Investigations on Three Species of Insects Recommended Schlettwein, C. H. G. 1985. Distribution and densities of Cyrtobagous singularis Hustache Semple, J. L. and I. W. Forno. 1987. Native parasitoids and pathogens attacking Samea multiplicalis Sharma, K. P. and P. K. Goel. 1986. Studies on decomposition of two species of Salvinia. Storrs, M. J. and M. H. Julien. 1996. Salvinia. A Handbook for the Integrated Control of Salvinia Taylor, M. F. J. 1984. The dependence of development and fecundity of Samea multiplicalis on early Taylor, M. F. J. 1988. Field measurement of the dependence of life history on plant nitrogen and Taylor, M. F. J. 1989. Compensation for variable dietary nitrogen by larvae of the salvinia moth. Taylor, M. F. J. and I. W. Forno. 1987. Oviposition preferences of the salvinia moth Samea Taylor, M. F. J. and D. P. A. Sands. 1986. Effects of aging and nutrition on the reproductive system of Thomas, P. A. 1980. Life-cycle studies on Paulinia acuminata (De Geer) (Orthoptera: Pauliniidae) Thomas, P. A. and P. M. Room. 1986. Taxonomy and control of Salvinia molesta. Nature 320: Williams, R. H. 1956. Salvinia auriculata Aubl.: the chemical eradication of a serious aquatic weed in [ Contents ] [ Previous ] [ Next ] |
|
| ||||
| |
The Bugwood Network and Forestry Images Image Archive and Database Systems The University of Georgia - Warnell School of Forestry and Natural Resources and College of Agricultural and Environmental Sciences - Dept. of Entomology Last updated on Thursday, July 19, 2018 at 01:38 PM Questions and/or comments to the | |||