|
|
In: Van Driesche, R., et al., 2002, Biological Control of Invasive Plants in the Eastern United States, USDA Forest Service Publication FHTET-2002-04, 413 p.
States with millions of dollars spent nationwide for control efforts (U.S. Congress, Office of Technology Assessment, 1993). In New York state alone, annual costs are estimated at $500,000.
Ecological damage. With the expansion of Eurasian watermilfoil, species diversity and abundance of native macrophytes declines (Smith and Barko, 1990; Madsen et al., 1991). Although in small tank experiments the native northern watermilfoil (Myriophyllum sibiricum Kom.) appears competitively superior, in the field, however, M. spicatum has replaced M. sibiricum over much of the temperate range of this species in North America (Valley and Newman, 1998). Suppression of native macrophytes is enhanced through the formation of a Eurasian watermilfoil canopy at the water surface, reducing light penetration. This canopy often forms early in the season before native macrophytes reach their maximum growth potential. Eurasian watermilfoil beds contain significantly fewer macroinvertebrates than native macrophyte communities (including benthic invertebrates) and have a reduced abundance of native fish species (Keast, 1984). Fish spawning areas and fish growth are reduced in lakes with large infestations of Eurasian watermilfoil. During certain times in winter, waterfowl forage extensively in milfoil beds in Alabama (McKnight and Hepp, 1998; Benedict and Hepp, 2000), although the same species largely avoid Eurasian watermilfoil in the Great Lakes (Knapton and Pauls, 1994). Extent of losses. Direct losses are difficult to quantify due to lack of data from long-term monitoring programs. Geographical Distribution From the initial points of introduction in the Northeast, M. spicatum has spread to 44 states and at least three Canadian provinces (Creed, 1998) and is now considered a major nuisance species throughout the Northeast, northern Midwest and Pacific Northwest of the United States (Couch and Nelson, 1985, White et al., 1993). The mode of dispersal is not completely understood, but M. spicatum can be spread short distances as fragments tangled on boats and trailers (Nichols and Shaw, 1986). Also, human activities, such as motor boating and mechanical weed harvesting, produce and distribute stem fragments allowing increased propagation (Nichols and Shaw, 1986). Long distance dispersal has been linked to the aquarium and aquatic nursery trade (Reed, 1977), and the species continues to expand its range in North America. Background Information On The Pest Plant Taxonomy Eurasian watermilfoil belongs to the watermilfoil family, Haloragaceae, which has two genera in the eastern United States, Myriophyllum L. (10 species, the watermilfoils) and Proserpinaca L. (two species, the mermaid-weeds) (Gleason and Cronquist, 1991). All species are hydrophytes with many finely divided leaves. All Haloragaceae species are herbs submersed in quiet waters or rooted on muddy shores. The similarity of the species has led to much confusion about species identity, and most species in the family cannot be separated using only individual specimens or ones without flowers. The date of introduction of M. spicatum to North America is debated and some authorities consider reports before 1940 as taxonomic misidentifications of M. sibiricum (= M. exalbescens Fern.) (Johnson et al., 1998). Myriophyllum spicatum is variable in appearance with long stems, and usually 12 to 21 leaflet pairs, which are limp when out of the water. In contrast, the very similar M. sibiricum usually has five to 10 leaflet pairs with leaflets that stay rigid when out of the water. Leaf morphology may be used to separate these two very similar species successfully (Gerber and Les, 1994). Plants often branch at the water surface (or in response to herbivore damage to apical meristems) and flowers are arranged on emersed spikes (associated with a dramatic shift in plant morphology). The flower spike bears whorls of female flowers basally and whorls of male flowers apically. Each female flower produces four small nutlike fruits (2 to 3 mm). Biology Eurasian watermilfoil occurs in ponds, lakes, and pools that vary from rather deep to very shallow (from more than 100 m to a few cm), and may be stagnant or slowly moving fresh to slightly brackish water (Spencer and Lekic, 1974). Plants overwinter rooted in the sediment and grow rapidly once favorable temperatures are reached. Flowering can occur in early summer and can continue for several months (Spencer and Lekic, 1974). Eurasian watermilfoil reproduces by seed, but fragmentation is the most likely mode of spread in the northern parts of the range in North America. Sexual reproduction appears unimportant in shaping population structure of Eurasian watermilfoil in Minnesota (Furnier and Mustaphi, 1992); however, significant germination is observed in Lake George in New York State (Hartleb et al., 1993). Seeds require high temperatures (above 14°C) for germination. Light is not considered a limiting factor, but increased sedimentation can greatly suppress germination (Hartleb et al., 1993). Under unfavorable conditions or when plants are attacked by herbivores, plants may not reach the water surface and do not flower. Fragmentation still allows populations to expand and colonize adjacent areas or reach more distant areas through the aquarium trade, transport in currents, or through recreational activities. Analysis of Related Native Plants in the Eastern United States The genus Myriophyllum belongs to the taxonomically isolated watermilfoil family, Haloragaceae, in the order Haloragales. The genus Myriophyllum is representative of this order; more distantly related species occur in the southern hemisphere. There has been much confusion about taxonomic status and identity of Myriophyllum species. Muenscher (1944) lists approximately 20 species of Myriophyllum in North America; Gleason and Cronquist (1991) list 10 species in the eastern United States. Three of the 10 Myriophyllum species mentioned by Gleason and Cronquist (1991) are species introduced to North America, including parrotfeather, Myriophyllum aquaticum (Velloso) Verdc, another invasive species. According to Gleason and Cronquist (1991), only one other genus within the Haloragaceae occurs in eastern North America represented by the two species of mermaid weeds: Proserpinaca palustris L. and Proserpinaca pectinata Lam. These species occur in the range of M. spicatum in North America and are widely distributed. Another North American genus, Hippuris (mare’s tail), has sometimes been included in the Haloragaceae but now is considered an independent family and outside the order Haloragales (Gleason and Cronquist, 1991). Another closely related order with many wetland plant species is the Myrtales, of which four families (Lythraceae, Thymelaceae, Onagraceae, and Melastomataceae) are native to much of North America. These groups also include invasive introduced species such as purple loosestrife (Lythrum salicaria L.) and water chestnut (Trapa natans L.) (see also the discussions of native species in these chapters). The closest relative to M. spicatum is northern milfoil M. sibiricum and the ranges of these species overlap widely in the temperate regions of North America. History of Biological Control Efforts in the Eastern United States For almost 30 years, overseas and domestic research has evaluated potential agents (insects and pathogens) for the biological control of Eurasian watermilfoil (Buckingham et al., 1981; Creed et al., 1992; Creed and Sheldon, 1993, 1995; Shearer, 1994; Sheldon and Creed 1995; Cofrancesco, 1998; Creed, 1998; Johnson, et al., 1998; Mazzei et al., 1999; Gross et al., 2001). Several species of insects have been identified feeding on and damaging Eurasian watermilfoil in North America (Batra, 1977; Buckingham and Bennett, 1981; MacRae et al., 1990; Creed and Sheldon, 1993). Some of these species appear to be native to North America and to have switched from their original hosts; others may have been accidentally introduced from Europe along with M. spicatum (Buckingham et al., 1981). Since 1963, the grass carp, Ctenopharyngodon idella (Cuvier and Valenciennes), has been released to suppress Eurasian watermilfoil and other nuisance aquatic plants in numerous sites within North America (Julien and Griffiths, 1998). Reports of recent declines in Eurasian milfoil abundance in some lakes in North America have been attributed to feeding damage of three herbivores, the midge Cricoptopus myriophylli Oliver, the weevil Euhrychiopsis lecontei Dietz, and the pyralid moth Acentria ephemerella Denis and Schiffermüller (syn.: Acentria nivea Olivier) (Painter and McCabe, 1988; MacRae et al., 1990; Creed and Sheldon, 1993; Sheldon and Creed, 1995; Newman et al., 1996, 1998; Johnson et al., 1998). Area of Origin of Weed Myriophyllum spicatum is native to Europe, Asia, and North Africa, where the species is widely distributed. Eurasian watermilfoil appears to have been accidentally introduced into North America sometime between the late 1800s and 1940 (Nichols and Mori, 1971; Couch and Nelson, 1985). The taxonomic difficulty in separating the native M. sibiricum and M. spicatum has made the exact date of introduction difficult to determine. A record of the accidentally introduced moth A. ephemerella from 1927 (Sheppard, 1945) suggests that M. spicatum arrived early in the 20th century or the moth was introduced with another plant species. Areas Surveyed for Natural Enemies Surveys for insects and pathogens with potential for biological control have been conducted throughout North America and in Europe (England, Denmark, former Yugoslavia) and Asia (Pakistan, Bangladesh, Korea, China) (Buckingham et al., 1981; Cofrancesco, 1998). Natural Enemies Found More than 20 insect species have been identified as feeding on M. spicatum overseas (Spencer and Lekic, 1974; Buckingham et al., 1981; Cofrancesco, 1998), but few were ever seriously investigated to determine their potential as biological control agents (Buckingham and Bennett, 1981; Buckingham and Ross, 1981), in part because many appear to lack necessary host specificity. Native or accidentally introduced insect herbivores with potential for control of Eurasian watermilfoil have received increasing attention over the past 30 years (Buckingham and Bennett, 1981; MacRae et al., 1990; Creed and Sheldon, 1995; Sheldon and Creed, 1995; Newman et al., 1996; Johnson et al., 1998; Creed, 2000; Gross et al., 2001). More recently, several pathogens have been evaluated as potential biological control agents (Smith et al., 1989; Shearer, 1994). Even though these pathogens, have occasionally shown promise in laboratory tests and been hypothesized to contribute to recent field declines of Eurasian watermilfoil (Shearer, 1994), none has been approved for field use. Among the insect species studied is the weevil Litodactylus leucogaster (Marsham), which appears to be a native North American species with a holarctic distribution (Buckingham and Bennett, 1981). Adults and larvae live on emersed flower spikes of various milfoil species. Since unfavorable growing conditions and herbivore attack often prevent M. spicatum from flowering, L. leucogaster appears to have little use in biological control programs. A second species, the aquatic midge Cricotopus myriophylli, has been reported as causing recent declines of Eurasian watermilfoil in British Columbia (MacRae et al., 1990). Laboratory experiments have demonstrated a negative impact of C. myriophylli on M. spicatum growth and biomass production (MacRae et al., 1990). This midge appears to be widely distributed in North America, from British Columbia to New York. However, taxonomic problems in the entire group make species identification difficult. Despite its wide distribution, and the potential it shows in the laboratory, little research has focused on quantifying the impact this herbivore may have on M. spicatum populations in the field. The midge does not appear to contribute significantly to declines in Eurasian watermilfoil performance, although field evidence is lacking. The most promising natural enemies meriting further investigation are the naturalized pyralid moth A. ephemerella and the native weevil E. lecontei. These species are discussed in detail in the Host Range Tests and Results section that follows. Host Range Tests and Results Formal host specificity screening has been conducted with A. ephemerella (Buckingham and Ross, 1981; Johnson et al., 1998; Gross et al., 2001) and to some extent with C. myriophylli (MacRae et al., 1990). Some initial screening tests have resulted in the immediate rejection of some candidate biological control agents (Buckingham et al., 1981). Acentria ephemerella. Tests with A. ephemerella specimens collected and tested in North America confirm reports from Europe (Berg, 1942) that the species has a preference for Eurasian watermilfoil, but is not monophagous. In the late 1970s, tests were conducted in quarantine using 20 test plant species with A. ephemerella specimens collected from New York state (Buckingham and Ross, 1981). In no-choice tests, larvae fed on almost all test plants offered but primarily on two species of mermaid weeds (P. pectinata and P. palustris), Hydrilla verticillata (L. fil) Royle, and several species of Potamogeton (pondweeds) (Buckingham and Ross, 1981). These results reduced the interest in using this generalist feeder as a potential biological control agent. Subsequent reports that A. ephemerella may be causing significant declines of Eurasian watermilfoil rekindled interest in the specificity of the species and Johnson et al. (1998) and Gross et al. (2001) conducted additional tests. In laboratory feeding assays, A. ephemerella larvae showed a clear preference for M. spicatum over Elodea canadensis Michx. (Gross et al., 2001). The lack of effect on the native E. canadensis was, in part, explained by differences in plant architecture. These results confirmed field observations that A. ephemerella has a feeding preference for M. spicatum, even though larvae can sometimes be observed feeding and overwintering on other submersed aquatic macrophytes. Larval feeding and cocoon/retreat construction on and near the apical meristem of Eurasian watermilfoil substantially reduces plant growth and often inhibits canopy formation. Similar feeding and cocoon/retreat construction on other co-occurring plant species attacked occasionally by A. ephemerella does not affect these species in this way due to differences in plant morphology and growth patterns (Johnson et al., 1998). No-choice feeding rate tests (Johnson et al., 1998) using seven species not previously tested by Buckingham and Ross (1981) showed that A. ephemerella feeds on other macrophytes but feeding rates are highest on Eurasian watermilfoil and pondweeds. On less preferred plant species, A. ephemerella larvae often consume plant tissue but fail to build cocoons and retreats resulting in reduced survival. Cricotopus myriophylli. In no-choice feeding and starvation trials using 12 different native macrophyte species (MacRae et al., 1990), larvae of the midge C. myriophylli fed to some extent on a native pondweed Potamogeton natans L., but regular feeding and construction of a case was only observed on M. exalbescens (= M. sibiricum). The strong feeding preference of C. myriophylli for M. spicatum and the absence of the species in areas outside the distribution of the introduced Eurasian watermilfoil suggests the species may be an accidental introduction (MacRae et al., 1990). Euhrychiopsis lecontei. The host specificity of the native weevil E. lecontei has not been investigated beyond comparing its preference for M. sibiricum versus the introduced M. spicatum (Solarz and Newman, 1996; Newman et al., 1997; Solarz and Newman, 2001). These tests showed that the host plant of the larvae influences performance and host plant choice of the progeny and that M. spicatum is usually the preferred host over M. sibiricum (Newman et al., 1997). Releases Made Releases of grass carp have been made throughout North America for control of aquatic nuisance species, including M. spicatum. The flower-feeding weevil L. leucogaster was released at a single location in Florida (Buckingham et al., 1981), but the species did not establish. Experimental releases of A. ephemerella were made in three New York state lakes (Hairston and Johnson, 2001), and in enclosure experiments. Several releases of E. lecontei were made throughout the region covered in this book including those conducted by lake associations and management agencies as well as researchers. The weevil is now commercially available. No other intended releases of insect herbivores have been made. Biology and Ecology of Key Natural Enemies Acentria ephemerella Denis and Schiffermüller (Lepidoptera: Pyralidae) The aquatic moth A. ephemerella is native to and widespread in Europe. The species is considered a generalist herbivore feeding on a variety of aquatic plants including Eurasian watermilfoil (Berg, 1942). The species was first reported in North America near Montreal in 1927 (Sheppard, 1945). Today, this moth has been found from Massachusetts to Iowa. Declining Eurasian watermilfoil populations in Ontario, Canada, and New York state have been associated with population explosions of A. ephemerella (Painter and McCabe 1988; Johnson et al., 1998; Gross et al., 2001).
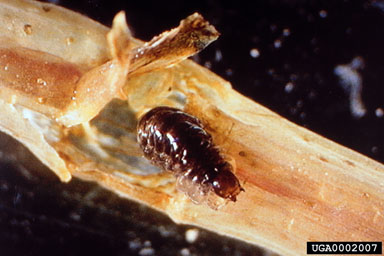
Larvae (0.25 mm in length) mine inside the small leaflets of Eurasian watermilfoil after hatching, until they are large enough to build resting shelters of leaflets and small branches that are attached to plant stems. The small size and cryptic behavior of the larvae make them difficult to detect and even trained observers often overlook them. Larvae disperse by crawling upward to feed on the apical meristem as they grow. Larvae are almost transparent, greenish caterpillars, usually with a visible dark green gut (ingested plant material). There are four or five larval instars and mature larvae are 10 or 12 mm long. When ready to pupate, they build a cocoon, often by removing the plant tip, adding it to a lower portion of the stem, and spinning a cocoon between them. Two generations of A. ephemerella per year have been observed in the field. Adults of the first generation emerge and lay eggs in June. Second generation larvae grow rapidly during the summer months and second generation adults emerge and lay eggs in late summer. Eggs laid late in summer hatch and larvae overwinter as various instars. Euhrychiopsis lecontei Dietz (Coleoptera: Curculionidae) The weevil E. lecontei, a native North American species and watermilfoil specialist, traditionally feeds on the native northern watermilfoil (Myriophyllum sibiricum). Although this weevil is widely distributed throughout the continent, little is known about its regional or local distribution (Jester et al., 1997). This weevil also feeds on Eurasian watermilfoil and may contribute to population declines. Weevils have reached extremely high densities at some locations (Newman et al., 1996; Johnson et al., 1998; Creed, 2000) occasionally as high as two or three adults or larvae per stem (Hairston and Johnson, 2001). Adult weevils overwinter in soil, leaf litter, and other cover close to shore and return to their summer habitat in lakes and ponds from overwintering sites in late April or early May depending on local weather conditions. Extensive shoreline development reduces suitable overwintering habitat and may limit winter survival of E. lecontei (Johnson et al., 1998). Weevils climb through existing vegetation or swim from shore to reach Eurasian watermilfoil beds. Fish predation may be an important weevil mortality factor (Sutter and Newman, 1997) and appears to greatly limit weevil populations in ponds (Hairston and Johnson, 2001). Adult weevils are small (3 mm in length) and dark-colored, with brownish black and yellowish stripes on the upper half of the body. These stripes fade to a lighter yellow-beige underneath (Fig. 6). Adults feed primarily on leaves and stems, causing a decrease in the amount of photosynthetic tissue. After mating, females may lay hundreds of eggs with an average of 1.9 eggs per day (Sheldon and O'Bryan, 1996), usually one egg per watermilfoil apical meristem. However, water temperatures need to have reached at least 10°C for eggs to be laid (Mazzei et al., 1999). Eggs (0.5 mm in diameter) are round and opaque yellow; larvae are creamy greenish or whitish tan with a dark purple-black head capsule. Larvae feed on the apical meristem and then mine the stem of the plant, consuming the cortex (Newman et al., 1996) (Fig. 7). As larvae near pupation, they turn purplish gray. Mature larvae prepare a pupation chamber further down the stem where they complete development. Each larva needs about 15 cm of Eurasian watermilfoil stem to complete development (Mazzei et al., 1999). Adults leave the pupal chamber to mate and lay eggs or swim ashore to overwinter (late summer generation).
Developmental rate is linearly related to temperature up to a threshold of 29°C with fastest development occurring above 25°C (Mazzei et al., 1999). Eggs hatch within 12 days at 15°C and within 4.2 days at 31°C. Complete development from egg to adult ranged from 16.6 days at 29°C to 61.7 days at 15°C with a lower developmental threshold of about 10°C (Mazzei et al., 1999). Complete development in the field typically requires 20 to 30 days. These data suggest that up to five generations per year are possible in the temperate parts of North America. However, in the field female weevils often appear to stop oviposition in mid August and prepare to overwinter, thus rarely allowing completion of even a fourth generation. The recent transfer of E. lecontei from the native northern watermilfoil, M. sibiricum, to the introduced M. spicatum did not result in performance declines on the new host (Newman et al., 1997). Data collected by Creed and Sheldon (1993) indicated that E. lecontei will feed on M. sibiricum when M. spicatum is not available but prefers M. spicatum when given a choice. In performance experiments, weevil survival on the two host plants did not differ significantly, but progeny of adults reared on Eurasian watermilfoil performed more poorly on northern watermilfoil than did progeny of adults reared on northern watermilfoil and development times were significantly longer on the native M. sibiricum (Solarz and Newman, 1996; Newman et al., 1997; Solarz and Newman, 2001). Evaluation of Project Outcomes Establishment and Spread of Agents Grass carp have been widely used to suppress many different aquatic nuisance plants. Regulations as to whether individual states allow grass carp releases vary widely. By 1991, grass carp had been introduced into 35 states, following initial releases in Arkansas and Alabama in 1963. In 14 states, releases of both fertile diploid and sterile triploid fish have been allowed; 15 states have allowed only sterile triploids, and 19 states have prohibited all importations (Julien and Griffiths, 1998). Problems associated with the use of grass carp include difficulties in guaranteeing sterility, escape of individuals into areas with native fish species and, in particular, lack of specificity. In fact, in choice-tests with different plant species, M. spicatum was the least preferred food for grass carp (Pine and Anderson, 1991) The voracious appetite of this fish has potential to eliminate much of the aquatic plant biomass, greatly reducing the native plants that serve as important food and shelter for invertebrates, fish, and waterfowl (McKnight and Hepp, 1995). The aquatic moth A. ephemerella now appears widespread from the east coast to Minnesota and Wisconsin (Johnson et al., 1998); however, the species is cryptic and probably often overlooked, particularly as early instars. Regional surveys in New York show that A. ephemerella is widespread and occasionally abundant in most lakes in New York State (Johnson et al., 1998). The midge C. myriophylli has been collected at the Cornell Experimental Ponds (Ithaca, New York) and at many other places throughout North America. Indeed, it may be present in many lakes, but taxonomic difficulties in identifying larvae have prevented an easy assessment of the species distribution (Johnson et al., 1998). The weevil E. lecontei occurred in 24 of 26 lakes surveyed in New York (Johnson et al., 1998) and appears widely distributed in North America. The species is now commercially available, but releases into lakes and ponds often only supplement existing (often much larger) populations (Hairston and Johnson, 2001) and therefore may not expand the range of the species. Several states require that the mass-reared individuals used for releases be offspring of locally collected weevils. Suppression of Target Weed In laboratory and in lake-enclosure experiments, A. ephemerella successfully controls Eurasian watermilfoil growth by destroying the apical meristem, reducing biomass and plant height, and preventing canopy formation. Long-term monitoring in Cayuga Lake, New York, showed a dramatic decline of Eurasian watermilfoil associated with large populations of A. ephemerella (Johnson et al., 1998; Gross et al., 2001). Ten years after the initial decline in Cayuga Lake, Eurasian watermilfoil biomass remained at very low levels with no canopy formation. The reduction in Eurasian watermilfoil constituted a 90% decline of the species in Cayuga Lake. As the Eurasian watermilfoil population declined, native plant species returned and they now dominate the plant community (Johnson et al., 1998; Gross et al., 2001). Prevention of canopy formation in Eurasian watermilfoil due to A. ephemerella herbivory was the most likely mechanism explaining the shift in dominance from M. spicatum to E. canadensis in Cayuga Lake (Gross et al., 2001). In the laboratory, in lake-enclosure experiments, and in the field, E. lecontei is capable of causing high levels of damage to the host plant (Creed and Sheldon, 1995; Sheldon and Creed, 1995; Newman et al., 1996; Hairston and Johnson, 2001; Newman and Biesboer, 2001). Certain declines of Eurasian watermilfoil have been associated with large populations of E. lecontei (Creed and Sheldon, 1995; Sheldon and Creed, 1995; Newman and Biesboer, 2001), and E. lecontei seems to be contributing to declines in Eurasian watermilfoil in some lakes in Vermont, Minnesota, and New York, (Creed and Sheldon, 1995; Sheldon and Creed, 1995; Newman et al., 1996; Newman and Biesboer, 2001) but not in others (Hairston and Johnson, 2001). High levels of seasonal weevil damage does not always translate into long-term declines in Eurasian watermilfoil populations (biomass or plant height) in lakes due to recovery of M. spicatum after adult weevils move to shore for overwintering (Hairston and Johnson, 2001). Recovery of Native Plant Communities One of the best documented long-term shifts in plant populations associated with feeding by aquatic herbivores occurred in Cayuga Lake in New York state, where standardized surveys for aquatic plant growth and associated herbivores have been conducted for more than a decade (Johnson et al., 1998). In years of greatest abundance of M. spicatum in Cayuga Lake, few other macrophytes were present and usually in low abundance. Since the discovery of A. ephemerella in 1991, Eurasian watermilfoil biomass has dramatically declined at both the north and south end of the lake to less than 10% of the original abundance. Although A. ephemerella is a generalist species found feeding on other macrophytes, the net result of the population explosion of this aquatic caterpillar has been a selective suppression of M. spicatum (Johnson et al., 1998). Although E. canadensis is a suitable (but not preferred) food plant in the field and in no-choice tests with A. ephemerella, E. canadensis is now the most abundant plant species in Cayuga Lake. Both the north and the south end exhibit a very different macrophyte community structure dominated by native plants such as Chara vulgaris L., Heteranthera dubia (Jacquin) MacMillan, and Vallisneria americana Michx. (Johnson et al., 1998). For most other locations, similar long-term data sets using standardized collection techniques and monitoring of aquatic herbivores are not available. Economic Benefits The successful control and further implementation of the biological control program in Cayuga Lake in New York has reduced herbicide use. In addition, lakeside communities that previously suffered economically due to reductions in boating, swimming, and fishing have rebounded. Recommendations for Future Work Areas of Needed Work At present, the focus in the biological control program against M. spicatum is on evaluation of impact and additional releases of E. lecontei (Creed, 2000). In addition, in some areas in New York state, the mass production of A. ephemerella, and development of procedures for its release and evaluation are underway (Hairston and Johnson, 2001). Evaluations of releases should be based on use of standardized monitoring protocols that provide baseline data on the macrophyte communities and their associated herbivores (such as described by Johnson et al., 1998). Monitoring should be of long duration and done by trained personnel to avoid overlooking the effects or presence of cryptic species such as A. ephemerella. More information is needed on the efficacy of making additional releases of E. lecontei, particularly releases where well established weevil populations already are present. An additional focus should be the continued evaluation, under field conditions, of the effects of A. ephemerella on native macrophyte communities. Results from Cayuga Lake show that the net result of feeding by this species can be an increase in native species diversity and abundance (Johnson et al., 1998; Gross et al., 2001). If this species is found to be beneficial despite its host-range, further releases and redistribution, as well as release and mass production procedures should be developed. Future Needs for Evaluation Studies The presence of several species of milfoil herbivores could allow research and monitoring programs to test the assumption of cumulative effects of herbivores. Combinations of agents are likely to be more destructive to plants than a single species alone (Harris, 1981; Malecki et al., 1993). However, even spatially separated herbivores can compete via their common host plant (Masters et al., 1993; Denno et al., 1995). Moreover, all species considered destructive on M. spicatum attack the apical meristems and are not spatially separated. Whether these potential competitive interactions have any influence on control of M. spicatum where E. lecontei, A. ephemerella, and C. myriophylli co-occur requires further study. Results from experiments and monitoring at Dryden Lake in New York state seem to suggest that large populations of E. lecontei can suppress A. ephemerella populations to very low levels by almost completely destroying Eurasian watermilfoil apical meristems and toppling the plants early in the season just as A. ephemerella larvae require them for cocoon building and adult emergence (Hairston and Johnson, 2001). However, M. spicatum is able to recover after the collapse and the weevils leave the lake to overwinter; yet A. ephemerella populations remain suppressed. Despite the mid-summer population crash, Eurasian watermilfoil continues to be the dominant plant species in Dryden Lake and E. lecontei is unable to control its population. Weevil densities in Dryden Lake (3.58 individuals per stem) are beyond the density of 1.5 weevils per stem that have been reported to be able to control Eurasian watermilfoil elsewhere (Newman et al., 1996; Sheldon 1997). The end result of this competitive interaction (at least in shallow Dryden Lake) of the two herbivores is that a species unable to permanently control the host plant, E. lecontei, is in fact reducing the impact of a successful biological control agent, A. ephemerella (Hairston and Johnson, 2001). This would be the first case where an unsuccessful biological control agent is able to interfere with control through a second species (Crawley, 1989; Lawton, 1990); however, these results may be restricted to shallow lakes (Johnson et al., 2000) and need further evaluation. Evidence from five lakes in New York suggests that A. ephemerella densities are negatively correlated with M. spicatum biomass, while E. lecontei densities are positively correlated with M. spicatum biomass (Johnson et al., 2000). In addition, A. ephemerella appears more abundant in larger, deeper lakes while E. lecontei populations are negatively correlated with lake mean depths (Johnson et al., 2000). Overall, moth densities are negatively correlated with weevil densities, further suggesting important interspecific competition (Johnson et al., 2000). However, we need a larger sample size to substantiate these results from New York. Mechanical harvesting or application of herbicides removes the host plants and any herbivores feeding on them. Such actions reduce populations of A. ephemerella, E. lecontei and C. myriophylli, and limit their potential to control Eurasian watermilfoil. In many areas where harvesting operations occur regularly, Eurasian watermilfoil beds are lush and show very little feeding damage, while Eurasian watermilfoil beds in the same lake that have remained unharvested collapse due to feeding by aquatic herbivores (Johnson, pers. obs.). The pressure by lake associations and recreational users for quick action and immediate reductions of aquatic macrophytes may prevent a longer lasting, ecologically sound, and less expensive biological control program from achieving its full potential. Additional demonstration projects and evaluation of the long-term effects of the available biological control agents are needed to withstand the constant pressure for a quick reduction in aquatic vegetation. References Aiken, S. G., P. R. Newroth, and I. Wile. 1979. The biology of Canadian weeds. 34. Myriophyllum Batra, S. W. 1977. Bionomics of the aquatic moth, Acentropus niveus (Oliver), a potential biological Benedict, Jr., R. J. and G. R. Hepp. 2000. Wintering waterbird use of two aquatic plant habitats in a Berg, K. 1942. Contributions to the biology of the aquatic moth Acentropus niveus (Oliver). Vidensk Boylen, C. W., L. W. Eichler, and J. W. Sutherland. 1996. Physical control of Eurasian watermilfoil in Buckingham, G. R. and B. M. Ross. 1981. Notes on the biology and host specificity of Acentria nivea Buckingham, G. R. and C. A. Bennett. 1981. Laboratory biology and behavior of Litodactylus Buckingham, G. R., C. A. Bennett, and B. M. Ross. 1981. Investigations of two insect species for Cofrancesco, Jr., A. F. 1998. Overview and future direction of biological control technology. Journal of Couch, R. W. and E. N. Nelson. 1985. Myriophyllum spicatum in North America, pp 8-18. In Crawley, M. J. 1989. The successes and failures of weed biocontrol using insects. Biocontrol News Creed, R. P. 1998. A biogeographic perspective on Eurasian watermilfoil declines: additional evidence Creed, R. P. 2000. Is there a new keystone species in North American lakes and rivers? Oikos 91: Creed, R. P. and S. P. Sheldon. 1993. The effect of feeding by a North American weevil, Creed, R. P. and S. P. Sheldon. 1995. Weevils and watermilfoil: Did a North American herbivore Creed, R. P., S. P. Sheldon, and D. M. Cheek. 1992. The effect of herbivore feeding on the buoyancy Denno, R. F., M. S. McClure, and J. M. Ott. 1995. Interspecific interactions in phytophagous insects: Furnier, G. R. and M. M. Mustaphi. 1992. Isozyme variation in Minnesota populations of Eurasian Gerber, D. T. and D. H. Les. 1994. Comparison of leaf morphology among submersed species of Gleason, H. A. and A. Cronquist. 1991. Manual of Vascular Plants of the Northeastern United States Gross, E. M., R. L. Johnson, and N. G. Hairston, Jr. 2001. Experimental evidence for changes in Hairston, Jr., N. G. and R. L. Johnson. 2001. Monitoring and evaluating the impacts of herbivorous Harris, P. 1981. Stress as a strategy in the biological control of weeds, pp. 333-340. In Papavizas, Hartleb, C. F., J. D. Madsen, and C. W. Boylen. 1993. Environmental factors affecting seed Jester, L. L., M. A. Bozek, S. S. Sheldon, and D. R. Helsel. 1997. New records for Euhrychiopsis Johnson, R. L., E. M. Gross, and N. G. Hairston Jr. 1998. Decline of the invasive submersed Johnson, R. L., P. J. Van Dusen, J. A. Toner, and N. G. Hairston, Jr. 2000. Eurasian watermilfoil Julien, M. H. and M.W. Griffiths. 1998. Biological Control of Weeds. A World Catalogue of Agents and Keast, A. 1984. The introduced aquatic macrophyte Myriophyllum spicatum as habitat for fish and Knapton, R. W. and K. Pauls. 1994. Fall food habits of American widgeon at Long Point, Lake Erie, Lawton, J. H. 1990. Biological control of plants: A review of generalizations, rules, and principles MacRae, I. V., N. N. Winchester, and R.A. Ring. 1990. Feeding activity and host preference of the Madsen, J. D., J. W. Sutherland, and C. W. Bloomfield. 1991. The decline of native vegetation under Malecki, R.A., B. Blossey, S. D. Hight, D. Schroeder, L. T. Kok, and J. R. Coulson. 1993. Biological Masters, G.J., V. K. Brown, and A. C. Gange. 1993. Plant mediated interactions between above- and Mazzei, K. C., R. M. Newman, A. Loos, and D. W. Ragsdale. 1999. Development rates of the native McKnight, S. K. and G. R. Hepp. 1995. Potential effects of grass carp herbivory on waterfowl foods. McKnight, S. K. and G. R. Hepp. 1998. Diet selectivity of gadwalls wintering in Alabama. Journal of Muenscher, W. C. 1944. Aquatic Plants of the United States. Cornell University Press, Ithaca, New Newman, R. M. and D. D. Biesboer. 2001. A decline of Eurasian watermilfoil in Minnesota associated Newman, R. M., K. L. Holmberg, D. D. Biesboer, and B. G. Penner. 1996. Effects of the potential Newman, R. M., D. C. Thompson, and D. B. Richman. 1998. Conservation strategies for the Newman, R. M., M. E. Borman, and S. W. Castro. 1997. Developmental performance of the weevil Nichols, S. A. and B. H. Shaw. 1986. Ecological life histories of three aquatic nuisance plants Nichols, S. A. and S. Mori. 1971. The littoral macrophyte vegetation of Lake Wingra. Wisconsin Painter, D. S. and K. J. McCabe. 1988. Investigation into the disappearance of Eurasian watermilfoil Pine, R. T. and L. W. J. Anderson. 1991. Plant preferences of triploid grass carp. Journal of Aquatic Reed, C. F. 1977. History and distribution of Eurasian watermilfoil in the United States and Canada. Shearer, J. F. 1994. Potential role of plant pathogens in declines of submersed macrophytes. Lake Sheldon, S. P. 1997. Investigations on the potential use of an aquatic weevil to control Eurasian Sheldon, S. P. and L. M. O'Bryan. 1996. Life History of the weevil Euhrychiopsis lecontei, a potential Sheldon, S. P. and R. P. Creed. 1995. Use of a native insect as a biological control for an introduced Sheppard, A. C. 1945. A new record for Canada (Lepidoptera). Canadian Entomologist 77: 55. Smith, C. S. and J. W. Barko. 1990. Ecology of Eurasian watermilfoil. Journal of Aquatic Plant Smith, C. S., S. J. Slade, J. H. Andrews, and R. F. Harris. 1989. Pathogenicity of the fungus, Solarz, S. L. and R. M. Newman. 1996. Oviposition specificity and behavior of the watermilfoil Solarz, S. L. and R. M. Newman. 2001. Variation in host plant preference and performance by the Spencer, N. R. and M. Lekic. 1974. Prospects for biological control of Eurasian watermilfoil. Weed Sutter, T. J. and R. M. Newman. 1997. Is predation by sunfish (Lepomi spp.) an8 important source of U. S. Congress, Office of Technology Assessment 1993. Harmful Non-Indigenous Species in the Valley, R. D. and R. M. Newman. 1998. Competitive interactions between Eurasian watermilfoil and White, D. J., E. Haber, and C. Keddy. 1993. Invasive Plants of Natural Habitats in Canada. Canadian [ Contents ] [ Previous ] [ Next ] | |||||||||||||
|
| ||||
| |
The Bugwood Network and Forestry Images Image Archive and Database Systems The University of Georgia - Warnell School of Forestry and Natural Resources and College of Agricultural and Environmental Sciences - Dept. of Entomology Last updated on Thursday, July 19, 2018 at 01:38 PM Questions and/or comments to the | |||